











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El género Callyntra Solier, 1836 incluye un conjunto de especies endémicas de Chile y la Argentina (Flores y Vidal, 2000a; Roig-Juñent, Flores, Ocampo y Smith, 2004). La distribución del género en Chile abarca toda la región de Chile central, y las especies habitan en ambas cordilleras (de los Andes y de la Costa) y en terrazas costeras (Flores y Vidal, 2000a,b). Luego de tener una historia taxonómica conflictiva, el género se estableció como grupo natural dentro de la tribu Nycteliini (Flores, 2000), lo que conllevó una revisión y revalidación del mismo (Flores y Vidal, 2000a) y se establecieron las relaciones filogenéticas entre las especies basadas en morfología (Flores y Vidal, 2000b). A pesar de ser un grupo taxonómicamente bien estudiado (ver Flores y Vidal, 2000a,b), recientemente ha existido una proliferación de trabajos que describen nuevas especies, como es el caso de C. hibrida Zúñiga-Reinoso y Cid-Arcos, 2013, C. zambranoi Aceituno y Guerrero, 2014, C. rufocastanea Aceituno y Guerrero, 2014, C. pehuenche Zúñiga-Reinoso y Cid-Arcos, 2015 y C. inconsiderata Zúñiga-Reinoso, 2016, teniendo en la actualidad 22 especies descritas.
Exploraciones recientes en la zona de Valparaíso han resultado en la recolección de especímenes que difieren de las especies previamente descritas, siendo morfológicamente más cercanos a la especie C. laticollis Philippi y Philippi, 1864. Por lo tanto, nuestro objetivo es describir una nueva especie para el género Callyntra con base en caracteres morfológicos externos y de los genitales.
Materiales y métodos
El material (n = 75) fue revisado bajo lupa estereoscópica y microscopio electrónico. Para ello, parte del material fue debidamente limpiado, montado en seco y etiquetado, mientras que otra parte fue conservado en alcohol al 99.8%. Para la descripción de la morfología externa se siguió a Flores y Vidal (2000a). Para establecer las diferencias entre las especies, según lo propuesto por Flores (1996), se examinaron los genitales de los machos, los cuales fueron comparados con material de colección de las especies morfológicamente cercanas. Para esto algunos ejemplares machos fueron ablandados con agua para la extracción de los genitales (n = 3) y posteriormente limpiados con una solución de KOH al 5% por 10 min. El edeago fue suspendido en solución de Hood en tubos Eppendorf de 0.2 ml. Los caracteres diagnósticos de la nueva especie se presentan conjuntamente con los C. laticollis, que es la especie morfológicamente más cercana. Los ejemplares examinados y asignados a la serie tipo se encuentran depositados en las siguientes colecciones: CPPP, Colección Particular de Pablo Pinto, Valparaíso, Chile; CPSL, Colección Particular de Sebastián Larrea, Viña del Mar, Chile; IADIZA, Instituto Argentino de Investigaciones de las Zonas Áridas, Mendoza, Argentina; IEUM, Colección Entomológica del Instituto de Entomología, Universidad Metropolitana de Ciencias de la Educación, Chile; MNNC, Museo Nacional de Historia Natural, Santiago, Chile; UCCC, Museo de Zoología, Universidad de Concepción, Concepción, Chile.
Descripción
Callyntra tortuosa Zúñiga-Reinoso, Pinto y Larrea n. sp. (figs. 1-2).

Figura 1. Paratipos en vista dorsal de Callyntra tortuosa de la localidad de Placilla: a) macho vista dorsal; b) hembra vista dorsal; c-d) detalles del meso-metaepisterno y esternitos abdominales de macho y hembra respectivamente. La escala representa 5 mm.
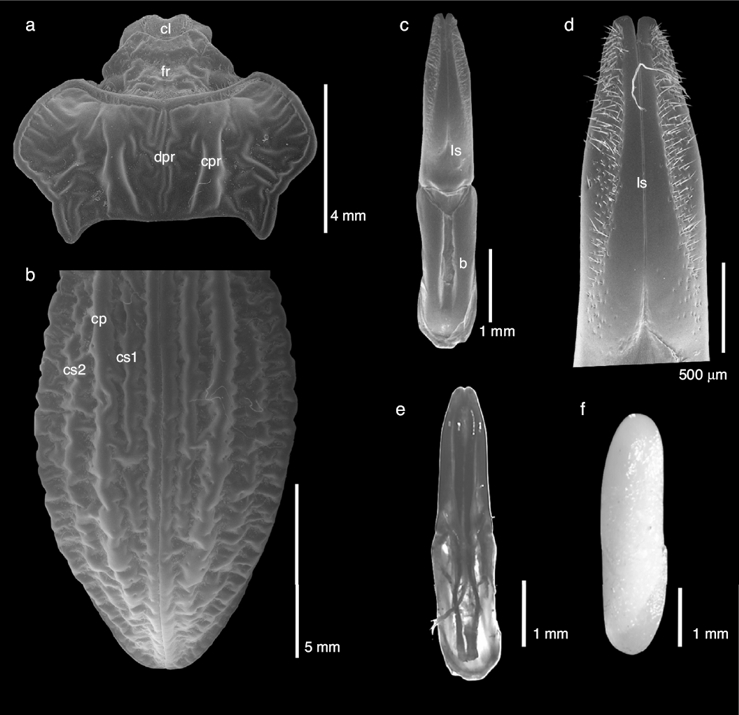
Figura 2. Estructuras aumentadas de Callyntra tortuosa. a) Cabeza y pronoto en vista dorsal; cl: clípeo, fr: frente, dpr: disco pronotal, cpr: carena pronotal. b) Élitros; cp: carena principal, cs: carena secundaria. c) Vista ventral de la genitalia; b: lámina basal del tegmen; ls: estilos laterales del tegmen. d) Detalles de los estilos laterales; ls: estilos laterales del tegmen. e) Vista dorsal de la genitalia. f) Huevo en vista lateral.
Holotipo. Cabeza. Coloración general negra (fig. 1). Labro con leves puntuaciones, escotado en V, con sedas largas en los bordes. Sedas cortas y esparcidas sobre la superficie. Clípeo subrugoso, con puntuación y sedas cortas escasas sobre la superficie (fig. 2a). Sutura clipeal no visible. Frente hundida, sin puntuación, con márgenes levantados. Arruga sobresaliente en centro de la frente, la divide en dos (fig. 2a). Mentón con fuerte puntuación y abundantes sedas largas. Antenas de coloración negra con abundante pilosidad blanca (fig. 1), teniendo los últimos 3 antenitos apariencia aterciopelada. La extensión de las antenas llega hasta ¾ del pronoto. Tercer antenito más largo que el primero y segundo; antenitos cuarto al sexto de proporciones semejantes; antenitos séptimo y octavo más pequeños que los anteriores y de medida semejante; noveno y décimo antenitos redondeados; antenito undécimo y final terminado en punta. Tórax. Pronoto plano, glabro y sin puntuación. Alcanza su ancho pronotal máximo en el centro de la estructura, debido a expansiones laterales redondeadas, y se estrecha abruptamente hacia la base del pronoto. Ángulos anteriores redondeados. Margen anterior sin puntuación, con abundante pilosidad corta que nace bajo este en dirección hacia la cabeza. Parte central del margen anterior levemente ensanchada, estrechándose hacia los costados. Margen lateral levantado, delgado, glabro y liso. Ángulos posteriores en punta, proyectados hacia atrás, por sobre los élitros. Disco con 2 carinas paralelas gruesas y muy levantadas, con presencia de escasa pero marcada rugosidad en el centro que corre casi longitudinal entre las carinas (fig. 2a). Abundantes arrugas desordenadas entre las carinas y el margen lateral. Proepisterno liso. Proesterno convexo, arrugado y presencia leve de sedas largas. Apófisis proesternal ovalada, levantada en la parte inicial y en declive hacia el ápice. Con puntuación leve y sedas esparcidas. Mesoventrito canaliculado en el centro, con leve puntuación por los bordes y algunas sedas largas. Metaventrito subrugoso con pocas sedas (fig. 1c). Élitros. Con presencia de sedas de color blanco diminutas y esparcidas en la mitad posterior de los élitros. Élitros con costilla prominente, con costillas secundarias presentes no enteras, presentando cortes pequeños a lo largo (fig. 2b). Sutura elitral levantada. Borde elitral levemente notorio y crenulado. Epipleura y pseudoepipleura glabra y lisa. Abdomen. Esternitos glabros y de coloración general negra brillante y semilisos (fig. 1c). Patas. Todas las patas de color negro. Coxas con pequeñas sedas en la base y con puntuación leve. Trocánter con abundante pilosidad. Fémures con abundante pilosidad de color blanco en su faz inferior, en el resto de la estructura con sedas esparcidas que en su faz superior. Tibias con pilosidad corta y gruesa. Tarsos con abundantes sedas. Cinco tarsitos en las patas protoráxicas, los 4 primeros de dimensiones semejantes, el quinto más largo, de tamaño 1:3. Cinco tarsitos en las patas mesotorácicas, el primero más largo que los tarsitos segundo al cuarto, quinto tarsito más largo que el primero. Cuatro tarsitos en patas metatorácicas, primero y cuarto más largos que segundo y tercero. Uñas simples en todas las patas.
Genitales. Edeago con los estilos del tegmen cóncavos, que se agudizan hacia el ápice (fig. 2c). Margen basal bisinuado, incompletamente esclerotizado, formando un espacio de forma aristada en el último tercio de los estilos (fig. 2d). Con sedas cortas y abundantes en el ápice que se van esparciendo hacia la base (fig. 2d). Lámina basal del tegmen con base subcuadrada (fig. 2c). El lóbulo medio tubular, la mitad de ancho que los estilos laterales de tegmen, con la abertura apical alargada, ápice redondeado y distalmente amplio (fig. 2e). Al comparar los genitales de C. tortuosa con los de C. laticollis especie morfológicamente cercana, se observa que esta estructura difiere considerablemente con C. laticollis, debido al menor tamaño de los genitales, a que posee los estilos del tegmen más robustos, cortos y curvados y la lámina basal del tegmen más corta.
Huevo. El huevo queda recubierto por una película de arcilla adherida al corion debido a un mucílago producido al momento de la oviposición. El huevo presenta forma elongada, de 4.2 mm de largo por 1 mm de ancho, polos redondeados, siendo levemente más ancho hacia la base. Exocorion de color blanquecino, aerópilas presentes y de apariencia lisa, pero están adornadas de leves y múltiples microrrugosidades (fig. 2f). El área micropilar en la base de la región apical del huevo, de forma circular, se encuentra ligeramente más oscura y sobresaliente.
Resumen taxonómico
Material examinado. Holotipo: Chile, Región de Valparaíso: Provincia Valparaíso: Placilla. 17. XII. 2012. col. P. Pinto. 1♂ (MNNC). Paratipos: Chile: Región de Valparaíso: Provincia Valparaíso: Placilla. 2. XII. 2009. col. S. Larrea. 1♀ (MNNC). 3. XII. 2009. col. P. Pinto S. 1♀ (CPPP). 19. IX. 2010. col. C. Pineda. 1♀ (MNNC). 7. X. 2010. col. C. Pineda. 1♂ (MNNC). 15. XI. 2010. col. P. Pinto. 1♀ (MNNC). 10. XI. 2010. col. C. Pineda. 1♀ (MNNC). 10. XII. 2010. col. P. Pinto S. 1♂ (CPPP). 01.X.2010. col. C. Pineda 1♀ (CPSL). 26. XI. 2012. col. S. Larrea. 4♀♀ (UCCC). 10. XII. 2012. col. S. Larrea. 1♀/1♂ (UCCC). 17. XII. 2012. col. P. Pinto. 5♀♀/5♂♂ (1 MNNC, 3 UCCC, 3IEUM, 3 CPPP). 15.X.2013. col. S. Larrea. 1♂ (IEUM). 28. X. 2014. col. S. Larrea. 3♂♂ (IEUM). 15. XII. 2013. col. P. Pinto S. 1♀ (CPPP). 12. XII. 2014. col. S. Larrea. 1♀/6♂♂ (2 IADIZA, 5 CPSL). 03. X. 2013. col. S. Larrea. 3♀♀/3♂♂ (CPSL). 16. XI. 2013. col. S. Larrea. 3♀♀/4♂♂ (CPSL). 12. XI. 2014. col. S. Larrea. 3♀♀ (CPSL). 05. XI. 2015. col. S. Larrea. 4♀♀/4♂♂ (CPSL). 25.XI.2015. col. S. Larrea. 2♀♀/4♂♂ (CPSL). 10. XII. 2015. col. S. Larrea. 2♂♂ (CPSL). 24. XII. 2015. col. S. Larrea. 3♀♀/2♂♂ (CPSL). Playa Ancha (?). 20. IX. 2010. col. C. Pineda. 1♂ (MNNC).
Distribución. Chile, V región, provincia de Valparaíso. Esto corresponde a la región entomofaunística Desierto de Coquimbo Sur (Peña, 1966) y al área de endemismo homónima (Flores y Vidal, 2000b), que en esta latitud abarca las zonas costeras.
Preferencia de hábitat. Todos los ejemplares fueron recolectados caminando en sitios abiertos altamente intervenidos, con remanentes de pastizal costero interior y bosque nativo del tipo esclerófilo. Actualmente el hábitat se encuentra con alto grado de intervención antrópica, siendo la flora nativa reemplazada por plantaciones de Pinus y Eucalyptus, además de presencia de microbasurales y proyectos inmobiliarios en las inmediaciones.
Biología. Esta especie prefiere los lugares abiertos con presencia de gramíneas, los individuos se encuentran activos durante la mañana y al atardecer, evitando las horas de mayor calor ocultándose bajo ramas, palos y entre la vegetación. Además se encuentra asociada a Sisyrinchium cuspidatum (Iridaceae), especie que utiliza como refugio y a su vez como alimento. Otra planta de la que se alimenta es Alstroemeria sp. Durante las primeras horas de la mañana pueden ser observadas bebiendo el agua en forma de rocío que queda depositada sobre briófitas y gramíneas. La oviposición fue realizada a unos 15 mm de profundidad aproximadamente.
Diagnosis y comentario. Los caracteres de C. laticollis se presentan entre paréntesis. Borde elitral leve (sobresaliente), proyecciones elitrales posteriores con punta aguzada (punta roma), carinas pronotales lisas (rugosas), prácticamente sin pilosidad en los élitros, solo aparece muy levemente el declive elitral (con pilosidad abundante en todo el élitro), proyección lateral del pronoto subangulosa semitriangular (proyección subangulosa redondeada), pronoto plano (convexo). Los caracteres únicos son las costillas elitrales secundarias irregulares e interrumpidas por cortes y el mesoventrito canaliculado en el centro (fig. 1c,d).
Etimología. El epíteto específico proviene del latín tortuosa (femenino, singular), que significa curvas y ondulaciones irregulares y en distintos sentidos, el cual hace referencia a las costillas elitrales secundarias irregulares e interrumpidas que presenta esta especie.
Variación intraespecífica. En general, existe una alta variación en las costillas secundarias entre individuos de la misma localidad, siendo prácticamente únicas en cada individuo. Lo mismo ocurre con las arrugas presentes en el pronoto. No se observaron diferencias morfológicas importantes entre los individuos de diferente localidad.
Dimorfismo sexual. Se presentan entre paréntesis los caracteres de las hembras. Pronoto plano (levemente convexo). Poca rugosidad entre las carinas pronotales y desordenadas (harta rugosidad y casi longitudinal). Élitros con pilosidad apenas notoria (con pilosidad notoria). Élitros angostos (élitros anchos). Tarsos largos (tarsos cortos). El largo promedio de los machos es 16.3 mm (DE ± 0.46), mientras que el largo promedio de las hembras es 17.87 mm (DE ± 0.5) (fig. 1).
Comentarios taxonómicos
Con base en las características morfológicas externas y del edeago, se propone que los individuos que se encuentran en la zona costera de la V región de Valparaíso pertenecen a la nueva especie Callyntra tortuosa. Esta especie sería morfológicamente cercana a C. laticollis, pero varios rasgos la diferencian de esta (ver diagnosis). En la descripción se detallan las características del edeago de C. tortuosa como lo propuesto por Flores (1996), debido a que es un carácter diagnóstico ampliamente utilizado en el género para diferenciar especies (Aceituno y Guerrero, 2014; Flores y Vidal, 2000a,b; Zúñiga-Reinoso y Cid-Arcos, 2013, 2015). En general, las hembras fueron más grandes que los machos, como lo documentado anteriormente por Zúñiga-Reinoso y Cid-Arcos (2013, 2015). Adicionalmente, se encontraron diferencias en algunas estructuras, como el tórax, élitros y coxas (para más detalles, ver descripción), como lo documentado para C. pehuenche por Zúñiga-Reinoso y Cid-Arcos (2015).
Este es el primer trabajo donde se describe el huevo para este género, el cual permite compararlo con el huevo descrito para Gyriosomus, otro género de Nycteliini que se puede diferenciar fácilmente porque presenta decoraciones hexagonales en el primer tercio apical (Pizarro-Araya, Jerez y Cepeda-Pizarro, 2005), mientras que el huevo de C. tortuosa presenta microrrugosidades en toda su superficie.
Callyntra tortuosa ha sido recolectada solo en 2 localidades (Playa Ancha es un registro dudoso) cercanas a la costa y ambas localidades están siendo destruidas y reemplazadas por proyectos inmobiliarios y plantaciones forestales, por lo que se estima conveniente que esta nueva especie endémica de las costas de Chile central sea incluida en algún estado de conservación, ya que, bajo los criterios del Reglamento de Clasificación de Especies del Ministerio de Medio Ambiente de Chile, hay argumentos suficientes para su protección (Jerez, Zúñiga-Reinoso, Muñoz-Escobar y Pizarro-Araya, 2015).

