











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La zona ribereña se define como el área de transición entre un río o corriente de agua y el ecosistema terrestre adyacente, incluyendo tanto el cauce como la llanura de inundación y ambientalmente influida por inundaciones periódicas (Malanson, 1993; Lowrance et al., 2000). Estas zonas tienen un gran valor ecológico por constituir una transición entre las zonas terrestre y acuática y formar un corredor biológico a través de diferentes regiones (Gregory et al., 1991; Malanson, 1993). De esta forma, las riberas constituyen uno de los sistemas ecológicos más complejos de la biósfera y uno de los más importantes para mantener la vitalidad del paisaje y sus ríos (Naiman y Décamps, 1990). Las riberas proporcionan funciones disímiles tales como: la reducción de la carga de nutrientes y de los sedimentos, la disminución de la erosión, la regulación de la temperatura del agua, constituyen el hábitat para una gran diversidad de organismos acuáticos y terrestres (Fernandes et al., 2011; Hubble et al., 2010; Meek et al., 2010; Scott et al., 2009; Yuan et al., 2009).
Las comunidades vegetales que se desarrollan en las zonas ribereñas se conocen como vegetación ribereña y debido a su proximidad e interacción, el cauce y el régimen de caudales están adaptados a las inundaciones periódicas proveyendo resistencia a la energía de los caudales (Tabacchi et al., 1996). Se trata de una vegetación con ciertas características de estructura y de funcionamiento que la diferencian de la vegetación presente en los bosques aledaños (Naiman et al., 2005). Existe un fuerte consenso de que este tipo de vegetación constituye un componente esencial de los ecosistemas fluviales (Ceccon, 2003; Naiman et al., 2005).
El grado de conocimiento del funcionamiento del ecosistema en los ríos y zonas ribereñas tropicales está poco desarrollado, aun cuando estos ríos constituyen más de 50% de la escorrentía terrestre y constituyen una fuente importante de aportación de carbono hacia los océanos, por lo que se necesita desarrollar estudios relacionados con este tema (Tiegs et al., 2019). Asimismo, el estudio de las comunidades de plantas ribereñas es importante ya que pueden utilizarse para fines de biomonitoreo de los ríos (Daniel et al., 2006; Mendoza-Cariño et al., 2014). Además de proporcionar criterios técnicos para implementar actividades de manejo dirigidos a conservar, restaurar y minimizar posibles impactos negativos en este tipo de vegetación (Camacho-Rico et al., 2006).
En la actualidad existen diferentes índices para valorar de forma rápida y sencilla la calidad de las riberas, la calidad de la vegetación y estimar el estado ecológico de los ríos. Entre ellos están el índice de hábitat fluvial (IHF; Pardo et al., 2002), el índice de calidad del bosque de ribera (QBR; Munné et al., 2003) y el índice de calidad ecológica de las riberas (RQI; González-del Tánago y García-de Jalón, 2011; González-del Tánago et al., 2006). El RQI permite determinar la calidad ecológica de las riberas a través de los procesos y la dinámica riparia, es decir, los cambios constantes debido a procesos continuos (Howard, 1992; Miller et al., 2003), en contraste con los otros métodos centrados casi exclusivamente en la valoración del estado de la cubierta vegetal, por lo que resulta un método más efectivo para determinar la condición ecológica en los sistemas ribereños (Barquín et al., 2011; González-del Tánago y García-de Jalón, 2011). Además, con este índice se pueden identificar los problemas existentes y definir las estrategias de gestión o manejo de acuerdo con la calificación obtenida del estado de la ribera (González-del Tánago y García-de Jalón, 2006).
La mayoría de las áreas ribereñas en el mundo, incluidas las de México, han sido degradadas por actividades humanas (de la Lanza-Espino et al., 2018; Demissie et al., 2013; Díaz-Pascacio et al., 2018; Gordon y Meentemeyer, 2006; Ortiz, 2019; Sunil et al., 2010). Estas actividades incluyen la agricultura, urbanización, minería a cielo abierto, extracción de agua, regulación del flujo, pastoreo de ganado y cambio de uso del suelo (Gantes et al., 2014). Acciones que a menudo resultan en la fragmentación de las galerías ribereñas y cambios en la estructura y la composición de su vegetación (Malmqvist y Rundle, 2002; Moreno-Jiménez et al., 2017; Ortiz, 2019; Richardson et al., 2007; Sirombra y Mesa, 2010).
La composición, la estructura y la diversidad de especies de la vegetación ribereña está determinada por factores ambientales como: las inundaciones periódicas o caída de árboles y los cambios climáticos altitudinales (Camacho-Rico et al., 2006; Lite et al., 2005). Además, estudios refieren que las actividades antrópicas y el tipo de uso del suelo adyacentes a las riberas influyen también sobre la estructura y la composición de las especies ribereñas (Aguiar y Ferreira, 2005; Gantes et al., 2014; Meek et al., 2010; Villarreal et al., 2012).
En México, la condición ecológica de los bosques ribereños a escala nacional no ha sido evaluada, sin embargo, a otras escalas se ha demostrado el alto grado de degradación y transformación que presentan en el país (Díaz-Pascacio et al., 2018; Meli et al., 2017; Ortiz, 2019; Rodríguez-Téllez et al., 2016). A pesar de la importancia que tiene la vegetación ribereña en la provisión de servicios ecosistémicos y en el mantenimiento de la biodiversidad, el impacto del tipo de cobertura sobre este tipo de vegetación está pobremente entendido (Méndez-Toribio et al., 2014). Por lo anterior, nuestro objetivo es evaluar el impacto del tipo de cobertura sobre la calidad de las riberas, la diversidad y la estructura de la vegetación leñosa ribereña en el río Margaritas al sureste de México.
Partimos de la hipótesis que el tipo de cobertura adyacente a la ribera del río predice su calidad al modificar atributos estructurales de la vegetación ribereña, particularmente la extensión en anchura, la cobertura y la distribución del corredor ribereño, la composición florística y la estructura de la vegetación. Predecimos que las selvas tendrán mejor calidad en sus atributos estructurales y funcionales, a diferencia de los tipos de vegetación desprovistos de cobertura arbórea. La baja calidad en las condiciones estructurales compromete, como consecuencia del impacto de diversas actividades antrópicas, la funcionalidad de las riberas, tales como la disminución de la diversidad de edades y la regeneración natural de especies arbóreas ribereñas, el estado de las orillas del cauce ordinario, la conectividad lateral de las riberas con las llanuras de inundación, la calidad del sustrato y la conectividad vertical.
La información obtenida será fundamental para el desarrollo de estrategias que contribuyan a la conservación, restauración y persistencia de la vegetación ribereña en ríos que están sometidos a la intensificación de actividades antrópicas hacia las partes bajas de sus cuencas hidrográficas. Al respecto, el río Margaritas en la sierra Madre de Chiapas, al sureste de México, es un ejemplo ideal porque en la parte alta de la cuenca se encuentra el área natural protegida Reserva de la Biosfera El Triunfo. Sin embargo, en la zona media y baja de esta cuenca se observa un cambio en el patrón de uso del suelo, intensificado por actividades productivas como la agricultura y la ganadería que han provocado la fragmentación y la eliminación de la vegetación ribereña (Tovilla, 2005).
Materiales y métodos
La cuenca del río Margaritas (93°07’57” - 92°59’06” O, 15°25’01” - 15°41’40” N) tiene una extensión de 19,475.81 ha, se encuentra en la región hidrológica 23 Costa de Chiapas (Conagua, 2009), en el municipio de Pijijiapan, Chiapas (Fig. 1). Se reporta para la cuenca una temperatura media de 27.6 °C y una precipitación anual de 2,596.5 mm (datos de la estación meteorológica 23018 de Conagua). Dentro de esta cuenca el área de estudio abarcó desde los 652 hasta los 56 m de altitud, específicamente en esta área el tipo de clima presente es cálido-húmedo (García, 1998). Otros datos específicos de variables climáticas y de disturbio por cada sitio de muestreo se pueden consultar en la Tabla 1. Las variables climáticas se obtuvieron de WorldClim versión 2.1 con una resolución espacial de 1 km2. Estas variables se generan por medio de interpolación de datos de estaciones meteorológicas y fueron desarrolladas para todo el mundo (Fick y Hijmans, 2017), mientras que las variables de disturbio se obtuvieron con la ayuda del software Qgis 3.10 (Qgis Equipo de desarrollo, 2019).

Figura 1 Localización de los tramos de muestreo en la cuenca del río Margaritas, Chiapas, México. Los nombres de los tramos se indican en la Tabla 1.
Tabla 1 Variables climáticas y de disturbio de los tramos de muestreo en el río Margaritas, Chiapas, México. Tipo de cobertura: S, selva; VSA, vegetación secundaria arbórea; SCA, sin cobertura arbórea. Variables climáticas: Prec, precipitación anual; Temp, temperatura promedio; Rad. Sol., promedio radiación solar. Variables de disturbio: Dist. Zon., distancia a zonas agropecuarias; Dist. Pob., distancia a poblados; Dist. Cam.: distancia a caminos.
| Tramo | Climáticas | Disturbio | ||||
|---|---|---|---|---|---|---|
| Prec. (mm) | Temp. (0C) | Rad. Sol. (kJ m-2 día-1) | Dist. Zon. (m) | Dist. Pob. (m) | Dist. Cam. (m) | |
|
|
|
|
|
|
|
|
En la zona más alta del área de estudio, la vegetación es selva mediana subperennifolia (Miranda y Hernández, 1963), mientras que en las partes media y baja es vegetación secundaria y pastizales, tanto cultivados como inducidos, que se originaron a partir de selva mediana subperennifolia y de la que aún se pueden observar parches aledaños a los sitios de estudio, selva alta siempreverde según Miranda (1952). El tipo de suelo del área muestreada es litosol (INEGI, 2002).
Se seleccionaron 13 tramos en el río principal y corrientes tributarias, ubicados a diferente altitud, con diferentes coberturas y uso del suelo representativos en la cuenca (fig. 1). Los tramos comprendieron un rango altitudinal entre 56 y 652 m snm y el número de orden del río varió desde 4 a 6 (Tabla 2). Un tramo, que es la unidad de muestreo empleada, es una longitud homogénea de un río donde se pueden extraer datos en una escala física (Frissell et al., 1986). Esta escala es apropiada para describir los patrones de distribución y la composición de especies de la vegetación, así como para evaluar los efectos a mediano y largo plazo de las actividades humanas en los ríos (Frissell et al., 1986; González-del Tánago y García-de Jalón, 2006).
Tabla 2 Características físicas de los tramos de muestreo en el río Margaritas, Chiapas, México, ordenados de acuerdo con el tipo de cobertura. S, Selva; VSA, vegetación secundaria arbórea; SCA, sin cobertura arbórea. Alt, Altitud; OC, orden del río; Pend, pendiente.
| Nombre del tramo | Alt. (m snm) | OC | Pend. (%) | Orientación de ladera | Cobertura | Tipo de cobertura |
|---|---|---|---|---|---|---|
| Loco 1 (S1) | 652 | 4 | 25.6 | Noroeste | Selva | S |
| La Mina1 (S2) | 612 | 4 | 19.0 | Noroeste | Selva | S |
| Ramón 1 (S3) | 460 | 4 | 24.9 | Noroeste | Selva | S |
| Loco 2 (VSA1) | 476 | 4 | 17.4 | Noroeste | Pastizal-acahual | VSA |
| La Mina 2 (VSA2) | 472 | 4 | 14.9 | Noroeste | Selva-acahual | VSA |
| Confluencia 1 (VSA3) | 347 | 6 | 13.7 | Oeste | Pastizal-selva | VSA |
| Confluencia 3 (VSA4) | 250 | 6 | 8.4 | Noroeste | Pastizal-acahual | VSA |
| Margaritas 4 (VSA5) | 168 | 6 | 7.6 | Oeste | Pastizal-acahual | VSA |
| Margaritas 1 (VSA6) | 128 | 6 | 2.1 | Oeste | Pastizal-acahual | VSA |
| Confluencia 2 (SCA1) | 319 | 5 | 6.8 | Este | Pastizal | SCA |
| Margaritas 3 (SCA2) | 68 | 6 | 1.9 | Norte | Pastizal | SCA |
| Ramón 2 (SCA3) | 386 | 4 | 11.8 | Oeste | Pastizal | SCA |
| Margaritas 2 (SCA4) | 56 | 6 | 1.5 | Suroeste | Pastizal | SCA |
Se definieron las clases de cobertura vegetal de acuerdo con la clasificación de Miranda y Hernández (1963). Para ello se utilizaron mapas de vegetación y uso del suelo para la cuenca (1:250000), procesados en ArcGis 10.2 y ERDAS, así como imágenes de satélite SPOT 5, sobre las cuales se superpusieron capas temáticas de vegetación del Instituto Nacional de Estadística y Geografía (INEGI, 2013). Los usos del suelo se clasificaron de acuerdo con los tipos de cultivo que se presentaron en la cuenca, como mango, palma africana, pastizales y agricultura (Hernández-Hernández, 2014).
La vegetación leñosa y la calidad de las riberas se evaluaron en los 13 tramos seleccionados, de 250 m de longitud y ancho variable, pues se representó el espacio ocupado por la vegetación ribereña desde la orilla del cauce según el tipo de valle. Para el análisis, los tramos fueron agrupados en 3 tipos de cobertura como se explica a continuación. Los tramos con vegetación de selva mediana subperennifolia fueron llamados: selva (S; n = 3); los tramos de vegetación secundaria (acahual-pastizal o pastizal-selva), se etiquetaron como vegetación secundaria arbórea (VSA; n = 6) y aquellos que tenían muy poca cobertura arbórea (pastizal o pastizal-uso urbano) fueron nombrados sin cobertura arbórea (SCA; n = 4) (Tabla 1).
En cada tramo se aplicó el índice de calidad ecológica de las riberas (RQI) propuesto por González-del Tánago et al. (2006) y González-del Tánago y García-de Jalón (2011), el cual permite evaluar la condición ecológica de las riberas, a partir de 7 atributos: 1) dimensión del espacio ripario ocupado por vegetación asociada al río (dimensión en anchura), en este atributo se identifica el ancho del corredor de vegetación riparia y se observa la presencia de restricciones debido a la influencia humana, además se toma en cuenta que el ancho puede reducirse naturalmente en valles confinados; 2) continuidad longitudinal de la vegetación leñosa (continuidad longitudinal), este atributo se estima con base en la continuidad del corredor de vegetación riparia, estimando la intensidad de la fragmentación a través del tamaño y frecuencia de áreas abiertas formadas por la acción humana y el uso del suelo; 3) composición y estructura de la vegetación ribereña (composición de vegetación), en el cual se identifican las especies, la densidad y la altura de los estratos de la vegetación riparia del tramo de estudio, basados en las condiciones de referencia o la vegetación potencial; 4) regeneración natural de las principales especies leñosas (regeneración natural), aquí se evalúa la diversidad de edades y regeneración de las principales especies leñosas ribereñas; 5) condición de las orillas (condición de las orillas), la evaluación de este atributo consiste en observar procesos de erosión y sedimentación locales relacionadas con la sinuosidad del cauce; 6) conectividad transversal del cauce con sus riberas y la llanura de inundación (conectividad transversal), este atributo evalúa la presencia de restricciones físicas que impidan la conexión hidrológica del cauce con su ribera durante períodos de inundación, y 7) conectividad vertical a través de la permeabilidad y el grado de alteración de los materiales y relieve de los suelos riparios (sustrato y conectividad vertical), en este atributo se observa el tipo y permeabilidad de los materiales que predominan en las riberas, y se evalúa su capacidad de infiltración y recarga de acuíferos. Los 3 primeros atributos determinan la estructura física del corredor ribereño, mientras que los 4 restantes están relacionados con el funcionamiento del sistema. La evaluación de los atributos de RQI se realizó de forma sistemática y el uso de la hoja de campo incluida en el protocolo del índice: los atributos 1, 2 y 3 se evaluaron en cada margen del río por separado, mientras que los atributos 4 al 7 se evaluaron conjuntamente en ambos márgenes (González-del Tánago y García-de Jalón, 2011). La calidad de las riberas se determinó a partir de la puntuación obtenida de la suma de los 7 atributos evaluados, con las siguientes categorías (González-del Tánago y García-de Jalón, 2011): muy bueno (150-130), bueno (129-100), moderado (99-70), pobre (69-40), malo (39-10) y muy malo (< 10).
En cada tramo se establecieron 3 parcelas de 100 m2 (10 × 10 m, ubicadas en paralelo a la corriente), a una distancia de 40 m entre ellas y seleccionadas al azar donde se registraron todos los árboles y arbustos de vegetación leñosa con diámetro del tronco a la altura del pecho ≥ 2.5 cm (DAP, medido a 1.30 m). De cada individuo se anotó el diámetro del tronco (cinta diamétrica) y la altura, estimada visualmente (siempre por un mismo observador para disminuir el sesgo de apreciación), empleándose una vara de longitud conocida, la cual se situó paralelo al tronco del árbol y se estimó cuantas veces se pudiera medir esa longitud en el árbol.
La determinación taxonómica de las especies vegetales se hizo con apoyo de especialistas en herbario, y la consulta de listados florísticos (Villaseñor, 2016). Se realizaron comparaciones con ejemplares digitalizados disponibles en las páginas electrónicas de JSTOR Global Plants (JSTOR, 2019), Global Biodiversity Information Facility (GBIF, 2019), el Portal de Datos Abiertos UNAM (IBUNAM, 2019), asimismo, se consultaron los registros del Sistema Nacional sobre Biodiversidad (Conabio, 2019). Para determinar las especies en alguna categoría de amenaza se consultaron la lista oficial de especies en riesgo mexicanas NOM-059-SEMARNAT-2010 (Semarnat, 2010) y la Lista Roja de la Unión Internacional para la Conservación de la Naturaleza (UICN, 2019).
El efecto del tipo de cobertura sobre la calidad de las riberas se determinó a través de un Anova unifactorial y las diferencias entre los grupos mediante la prueba de rangos múltiples de Tukey, con el uso del programa R (R Development Core Team, 2018). Los datos cumplieron los supuestos de normalidad e igualdad de varianza una vez realizadas las pruebas de Shapiro-Wilk y Bartlett, respectivamente.
Se realizaron análisis de la diversidad con base en la perspectiva de la diversidad verdadera representada por los números de Hill (q D). Estos números son una medida del grado de distribución de las abundancias relativas entre las especies, donde 0D es la riqueza o número total de especies de la muestra; 1D, es el número efectivo de especies abundantes (o comunes) y fue calculado a partir del exponencial del índice de entropía de Shannon; mientras que 2D, es el número efectivo de especies muy abundantes (o dominantes) en la muestra, y se calculó mediante el inverso del índice de concentración de Simpson (Chao et al., 2014). También se obtuvieron los porcentajes de representatividad de las especies esperadas con los estimadores de diversidad ACE (Abundance-based coverage estimator, Chao y Lee, 1992) y Chao 1 (Chao, 1984; Chao y Lee, 1992). Para el cálculo de la diversidad beta, se utilizó el índice Chao-Jaccard, basado en la abundancia, pues se ha demostrado que tiene un sesgo menor que otros índices cuando falta una proporción sustancial de especies en la muestra (Chao et al., 2005). Considerando que el tamaño de muestra entre los tipos de cobertura fue diferente, los índices de diversidad se obtuvieron mediante un análisis de Rarefacción individual con el programa Past 3.21 (Hammer et al., 2012), posteriormente se compararon los promedios en el mismo programa a través de un ANOVA unifactorial, y las diferencias entre los grupos mediante la prueba de rangos múltiples de Tukey.
Para identificar las especies de plantas dominantes por cada tipo de cobertura se construyeron curvas de rango-abundancia (Whittaker, 1965). La estructura se analizó por medio del área basal (m2 ha-1) de cada especie por sitio, el promedio de la altura de los árboles y el valor de importancia relativa por especie (VIR), obtenido de la sumatoria de los valores relativos de la abundancia (número de individuos), la frecuencia y la dominancia (área basal) (Mostacedo y Fredericksen, 2000). La composición de especies se comparó entre los tipos de cobertura (S,VSA y SCA) a través de un análisis Permanova usando la abundancia de las especies como variable dependiente con 9,999 permutaciones y con el uso de una matriz de distancia de Bray-Curtis. Cuando se detectaron diferencias se realizó un análisis Simper para determinar la contribución de las especies a esas diferencias.
Para analizar si existe relación entre la composición de especies y la diversidad con otras variables topográficas: altitud, pendiente de los tramos de muestreo y orientación de ladera (Tabla 2); climáticas: precipitación, temperatura, radiación solar (Tabla 1); y de disturbio: distancia a asentamientos humanos, zonas agropecuarias, caminos (Tabla 2), se realizó un análisis de escalamiento no métrico multidimensional, NMDS (por sus siglas en inglés), con el uso de una matriz de distancia de Bray-Curtis en el programa R (R Development Core Team, 2018). El análisis NMDS constituye la forma más robusta de ordenación para la detección de patrones ecológicos (Minchin, 1987).
Resultados
Los valores del RQI variaron de 36 a 136 entre los sitios (Tabla 3). Los valores más altos se obtuvieron en los tramos con selva adyacentes (S), mientras que los más bajos se obtuvieron para los tramos sin cobertura arbórea (SCA; Fig. 2). El Anova del RQI mostró diferencias significativas entre los 3 tipos de cobertura (F (2,10) = 30.37, p < 0.0001) y la prueba de Tukey indicó que los tramos con S tuvieron mayor RQI que los tramos de vegetación secundaria arbórea (VSA) (p < 0.005) y mayor que los tramos SCA (p < 0.0001).
Tabla 3 Condición de las riberas en los tramos con diferente cobertura en el río Margaritas, Chiapas, México. Tipo de cobertura: S, selva; VSA, vegetación secundaria arbórea; SCA, sin cobertura arbórea.
| Tramos | Uso del suelo adyacente a las riberas | Tipo de cobertura | Valor del RQI | Estado de las riberas |
| S1 | Vegetación | S | 136 | Muy bueno |
| S2 | Vegetación | S | 135 | Muy bueno |
| S3 | Vegetación | S | 112 | Bueno |
| VSA1 | Ganadería | VSA | 81 | Moderado |
| VSA2 | Vivero forestal | VSA | 101 | Bueno |
| VSA3 | Ganadería, camino | VSA | 110 | Bueno |
| VSA4 | Ganadería | VSA | 76 | Moderado |
| VSA5 | Área recreacional, camino | VSA | 76 | Moderado |
| VSA6 | Camino | VSA | 88 | Moderado |
| SCA1 | Poblado, camino | SCA | 53 | Pobre |
| SCA2 | Poblado, ganadería, área recreacional, camino, puente de autopista, bordos para evitar la inundación | SCA | 36 | Malo |
| SCA3 | Poblado, ganadería, | SCA | 63 | Pobre |
| SCA4 | Poblado, ganadería, puente de autopista, bordos para evitar la inundación | SCA | 40 | Pobre |
Puntajes RQI: Muy bueno: valores de 150-130; bueno: 129-100; moderado: 99-70; pobre: 69-40; malo: 39-10 y muy malo: < 10.
Los nombres de los tramos se indican en la Tabla 1.

Figura 2 Diagrama de caja del RQI por tipo de cobertura en los tramos muestreados del río Margaritas, Chiapas, México.
Los atributos del RQI variaron de malas a muy buenas condiciones (Fig. 3). Para los tramos en general, la regeneración natural, la composición de vegetación, así como la continuidad longitudinal fueron los atributos del RQI mayormente evaluados en las condiciones de pobre y malo, mientras que la conectividad transversal fue el atributo con mayor cantidad de tramos evaluados en las condiciones de moderados a muy buenos.
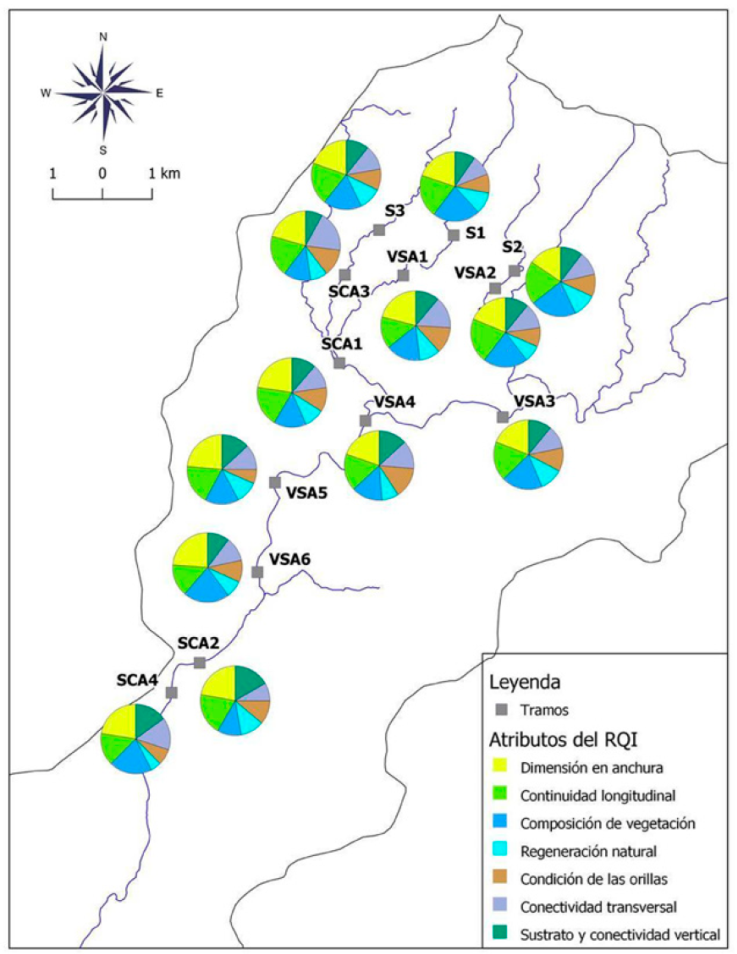
Figura 3 Gráficos de pastel de los atributos del RQI en los tramos muestreados del río Margaritas, Chiapas, México. Los nombres de los tramos se indican en la Tabla 1. El nombre para la etiqueta de cada atributo del RQI se describe en la sección de los métodos.
Los tramos con selva, ubicados a mayor altitud, con menor accesibilidad (i.e., alejados de poblados y caminos, con pendientes más inclinadas) y alejados de zonas agropecuarias presentaron los valores más elevados del índice y, por tanto, mejor calidad de las riberas. En estos tramos todos los atributos presentaron buenas y muy buenas condiciones, observándose riberas en buen estado de conservación, sin restricciones para el desarrollo de la vegetación ribereña y con abundante regeneración natural debido a la ausencia de barreras físicas naturales o provocadas por la intervención humana. No se observaron especies invasoras. En estos tramos, no existen restricciones físicas visibles (i.e., bordos y canalizaciones) que impidan la conectividad transversal y vertical del cauce con los restantes elementos del sistema fluvial (i.e., riberas, llanura de inundación y el medio hiporreico). Se observaron orillas estables sin indicio de erosión.
Por otro lado, los tramos SCA fueron evaluados mayormente con atributos pobres, moderados y de malas condiciones ecológicas (Fig. 3). En estos tramos se observó la disminución de las dimensiones del espacio ripario ocupado por vegetación. También se observó la presencia de ganadería bovina y de asentamientos humanos adyacentes a las riberas (Tabla 3). Se observó un alto grado de fragmentación de la vegetación, la cual fue sustituida con pastos para la alimentación del ganado. Se identificó la disminución de la regeneración natural de las principales especies leñosas y una disminución de la riqueza de especies, hubo indicios de erosión de las orillas, así como el ensanchamiento del canal y la disminución de la conectividad transversal del cauce con la llanura de inundación, debido a la construcción de viviendas, caminos y bordos para la protección contra la inundación. Se observó compactación del suelo y la modificación del relieve y los materiales naturales del terreno, lo cual conduce a la disminución de la conectividad vertical.
En los 13 tramos muestreados se registraron un total de 1,099 individuos, pertenecientes a 34 familias, 71 géneros y 90 especies. Una es especie exótica o introducida (Apéndice 1). En los tramos con VSA se identificó un total de 54 especies, seguido de S con 51 y los tramos SCA con 33 (Apéndice 1).
La familia más abundante fue Fabaceae (37%), seguida por Salicaceae (9%) y Urticaceae (6%). La especie con mayor cantidad de individuos (103) fue Vachellia cornigera (L.) Seigler y Ebinger. De acuerdo con la lista roja de la UICN (2019) las especies Cymbopetalum baillonii R.E. Fries, encontrada en tramos VSA, y Sapium macrocarpum Müll.Arg. que fue encontrada en tramos S, están consideradas como vulnerables (Apéndice 1). De acuerdo con la lista oficial de especies amenazadas de México, Croton guatemalensis Lotsy, observada en los tramos con S, está sujeta a protección especial, Licania arborea Seem. (observada en SCA) está considerada como amenazada y Vatairea lundellii (Standl.) Killip (observada en VSA) está catalogada en peligro de extinción (Apéndice 1).
Los tramos ribereños con S y VSA presentaron una distribución de serie normal logarítmica, con un pequeño número de especies abundantes y una gran proporción de especies raras. En los sitios SCA, si bien las comunidades muestran mayores niveles de uniformidad, el modelo no llega a ser de vara partida (Fig. 4). Las 3 especies de mayor abundancia relativa por cada tipo de cobertura fueron: en S, Justicia aurea Schltdl., Acalypha macrostachya Jacq. y Vachellia cornigera; en VSA, V. cornigera, Salix humboldtiana Willd. y Andira inermis (W. Wright) DC.; en los tramos SCA, Samanea saman (Jacq.) Merr., A. inermis e Inga vera Willd. (Fig. 4).

Figura 4 Curvas de rango-abundancia por tipo de cobertura de los tramos muestreados en el río Margaritas, Chiapas. La clave para las especies es: J_aur: Justicia aurea, A_mac: Acalypha macrostachya, V_cor: Vachellia cornigera, S_hum: Salix humboldtiana, A_ine: Andira inermis, S_sam: Samanea saman, I_ver: Inga vera, I_fag: Inga fagifolia, F_gla: Ficus glabrata, B_ali: Brosimum alicastrum, F_gol: Ficus goldmanii, B_sim: Bursera simaruba, B_cra: Byrsonima crassifolia, A_nio: Albizia niopoides, I_jin: Inga jinicuil, C_pol: Couepia poliandra.
La composición de especies mostró diferencias significativas entre los 3 tipos de coberturas (Permanova, F = 1.67, p = 0.03) siendo la comparación entre los tramos S y SCA la que resultó ser diferente (Permanova, F = 2.96, p = 0.03). De igual forma el análisis Simper indicó que la mayor disimilitud en la abundancia y composición de especies ocurrió entre los tramos de S y SCA (93%), siendo J. aurea, S. saman y A. inermis las especies que más contribuyeron con las diferencias encontradas. En el Apéndice 1, se muestra la composición de especies por cada tipo de cobertura. Los 3 tipos de coberturas comparten 9 especies, mientras que 18 especies se encontraron exclusivamente en S, 13 en tramos VSA y 5 especies en SCA.
El efecto del tipo de cobertura sobre la diversidad y estructura de la vegetación ribereña en los tramos muestreados se indica en la Tabla 4. El número de individuos fue mayor en los tramos con S y VSA que en tramos SCA. Similar tendencia mostró el área basal, donde se observó que en los tramos SCA se reporta la mitad del área basal respecto a los tramos S, así como la disminución del diámetro del tronco a la altura de 1.30 m.
Tabla 4 Efecto del tipo de cobertura sobre la estructura y la diversidad de la vegetación ribereña en el río Margaritas, Chiapas, México. Tipo de cobertura: S, selva; VSA, vegetación secundaria arbórea; SCA, sin cobertura arbórea.
| Variables | Tipo de cobertura | |||
|---|---|---|---|---|
| S | VSA | SCA | F | |
| Estructura | ||||
| Número de individuos/ha | 2,880 (311.0)a | 2,091.6 (154.4)b | 1,771.2 (151.6)c | 7.59** |
| Área basal (m2 ha-1) | 41.4 (0.15)a | 30.2 (2.38)b | 19.3 (1.72)c | 20.5*** |
| Altura (m) | 6.23 (0.93) | 6.24 (0.68) | 6.70 (0.54) | 0.15 n.s |
| Diversidad α | ||||
| Riqueza de especies (0D) | 33 (4.17)a | 32 (3.02)a | 19 (2.02)b | 6.10** |
| Número de especies comunes (1D) | 20 (2.71)a | 18 (1.36)a | 12 (1.08)b | 7.84** |
| Número de especies dominantes (2D) | 14 (1.18)a | 12 (0.78)a | 8 (0.70)b | 8.17** |
| ACE (Abundance Coverage-based) | 49 (4.59)a | 39 (3.32)a | 24 (2.42)b | 11.23*** |
| Chao 1 | 56 (5.0)a | 39 (3.17)b | 26 (2.93)c | 14.01*** |
| Diversidad β | ||||
| Chao-Jaccard Abundance-based | 0.35 (0.02) | 0.16 (0.05) | 0.19 (0.09) | 2.08 n.s |
Se indica la media (± error estándar), así como las diferencias obtenidas mediante el Anova unifactorial. * = p < 0.05, ** = p < 0.01, *** = p < 0.0001), n.s = no diferencia significativa. Diferentes letras denotan diferencias significativas entre los tipos de uso del suelo en las variables evaluadas (obtenidas con la prueba Tukey, α = 0.05). Los grados de libertad fueron: variables de estructura: g.l. 2,12; variables de diversidad α: g.l. 2,38; variables de diversidad β: g.l. 2,12).
La diversidad verdadera y las estimaciones de la diversidad alfa (ACE y Chao 1), fueron claramente afectadas por el tipo de cobertura (Tabla 4). La diversidad verdadera fue mayor en los tramos S, seguidos de los VSA y los tramos SCA. Con los valores de la riqueza verdadera y de los estimadores se obtuvieron los porcentajes de representatividad de las especies esperadas. Con el estimador (ACE), la representatividad de las especies esperadas fue de 67% para S, 82% los tramos VSA y el 79% para SCA. Con el segundo estimador (Chao 1), los porcentajes fueron, 59% en S, 82% en VSA y 73% en SCA. Los estimadores de diversidad mostraron que aún faltan especies por ser detectadas (p.ej., aproximadamente 16 especies en los tramos S con el estimador ACE). Finalmente, la comparación de la diversidad beta y la altura de las plantas no mostraron diferencias significativas.
Las 10 especies estructuralmente más importantes en la vegetación ribereña muestreada del río Margaritas (Apéndice 2) fueron: Andira inermis (VIR 23.78/300), Salix humboldtiana (23.26), Vachellia cornigera (18.22), Ficus pertusa L.f. (14.31), Samanea saman (13.83), Guazuma ulmifolia Lam. (9.01), Ficus maxima Mill. (8.71), Trema micrantha (8.70), Inga vera (8.31) y Justicia aurea (7.07). En el Apéndice 3 se muestran por cada tramo las 2 especies más importantes de acuerdo con el VIR obtenido. Dentro de este grupo destacan las especies V. cornigera y A. inermis las cuales obtuvieron cada una el mayor VIR en 3 de los 13 tramos. La especie V. cornigera en S3, VSA6 y VSA4, mientras que A. inermis tuvo mayor importancia en VSA5, VSA1 y SCA3. Otra especie que destacó por su VIR fue S. saman en los tramos SCA2 y SCA4 (Apéndice 3).
El NMDS mostró que los sitios se separan según el tipo de cobertura y las variables ambientales: la distancia a zonas agropecuarias (r2 = 0.78), la pendiente (r2 = 0.65) y la altitud (r2 = 0.68), en el eje 1 (correlacionados con éste de forma negativa) y la temperatura (r2 = 0.786) en el eje 2 (correlacionada de forma positiva) (Fig. 5). La gráfica muestra que los sitios de S están más alejados de zonas agropecuarias, tienen mayores pendientes, se encuentran a mayor altitud y presentan temperaturas más bajas.

Figura 5 Diagrama de ordenación de los 13 tramos de muestreo producidos por el análisis NMDS. Tipo de cobertura: S, selva; VSA, vegetación secundaria arbórea; SCA, sin cobertura arbórea. Se representan las variables ambientales significativas p <0.05. Dist. Zon, distancia a zonas agropecuarias, Pend, pendiente; Temp, temperatura. La altitud está representada por las cotas de la altitud. Los nombres de las especies que aparecen en la gráfica están abreviados con las 3 primeras letras del género y de la especie; los nombres completos se pueden consultar en el Apéndice 1.
Discusión
Acorde con la hipótesis planteada nuestros resultados indicaron que el tipo de cobertura adyacente influye en la condición de la zona ribereña al modificar atributos estructurales de la vegetación ribereña como la extensión en anchura, la cobertura y distribución del corredor ribereño como lo han planteado otros autores (Aguiar y Ferreira, 2005; Díaz-Pascacio et al., 2018; Kutschker et al., 2009; Meek et al., 2010; Méndez-Toribio et al., 2014). En general, la condición de la zona ribereña en los tramos evaluados resultó clasificado en un rango de muy buena a una mala condición, observándose que los tramos SCA, con mayor accesibilidad (i.e., cercanos a poblados y caminos, con pendientes menos inclinadas), y con la presencia de ganadería como uso de suelo adyacente a las riberas, fueron los que resultaron con los valores más bajos de RQI.
A lo largo de los tramos SCA se encontraron distribuidos los principales poblados y aledaño a éstos se encontraron las áreas en las que se desarrolla la actividad ganadera con mayor intensidad, lo cual explica en gran medida los bajos valores de RQI obtenidos para estos tramos. El pastoreo de ganado es una actividad que impacta negativamente la calidad de las riberas (Kutschker et al., 2009), pues el ganado en las zonas ribereñas incrementa la compactación del suelo, reduciendo la infiltración, degradando los márgenes de los cursos de agua y disminuyendo los niveles freáticos (Romero et al., 2014). Además, los suelos de las riberas utilizadas como abrevaderos son particularmente afectados por el ganado, aun cuando la carga sea baja o moderada (Gantes et al., 2014) ya que modifica las propiedades físicas del suelo, como el aumento de la densidad aparente y la pérdida de capacidad para la retención de agua, cambios que facilitan los procesos erosivos (Taboada y Micucci, 2009).
Los resultados mostraron que en las zonas más altas de la cuenca es donde se encuentran las mejores condiciones ecológicas del bosque ripario, similar al patrón observado en México para el río La Sauceda (Rodríguez-Téllez et al., 2016). En otros países también se ha observado este patrón, por ejemplo en el río Maullín en Chile, donde las mayores calidades del bosque se registraron en la parte alta de los ríos (Fernández et al., 2009); mientras que Suárez y Vidal-Abarca (2000) en el río Segura en España, observaron que los sitios con las mejores condiciones son los pertenecientes a las cabeceras de los mismos. Por su parte, Sirombra y Mesa (2012) reportaron una correlación positiva y significativa entre los valores de la calidad de las riberas y la altitud en ríos Andinos subtropicales, indicando que la más alta calidad corresponde a los sitios con mayores altitudes. La accesibilidad a los tramos está determinada por este gradiente de altitud, factor que explica el mayor impacto antropogénico en sitios de menor altura (Sirombra y Mesa, 2010).
Estos resultados también se explican por la existencia en la parte alta de la cuenca del río Margaritas de un área protegida, la Reserva de la Biosfera El Triunfo, con importantes valores naturales con un buen nivel de conservación y políticas de protección ambiental (Carabias et al., 1998; López et al., 2011). Sin embargo, hacia la parte media y baja de esta cuenca el desarrollo de actividades agrícolas y ganaderas ha propiciado un importante cambio de uso del suelo modificando el tipo de cobertura existente, con el predominio de grandes áreas de pastizales y vegetación secundaria, también conocida como acahual (Miranda y Hernández, 1963). En el estado de Chiapas este patrón de intensidad de uso del suelo, el cual ha conducido a la pérdida de más de 95% de la cobertura vegetal original, es característico de otras cuencas hidrográficas como la del río Cacaluta y la cuenca del río Vado Ancho (Gálvez-Marroquín, 2017; Hernández-Hernández, 2014; Tovilla, 2005). En esta última cuenca se reportó la pérdida, en el período de 2003 al 2015, de 860 ha de selva que se convirtieron principalmente en pastizales y vegetación secundaria (Gálvez-Marroquín, 2017). De igual forma, fue reportado en Guerrero, para la cuenca del río Coyuquilla, que la cubierta vegetal ha sido reducida en 38% producto de la ampliación de las áreas agrícolas y el establecimiento de pastizales inducidos para ganado (Osorio et al., 2015). En otras cuencas costeras de México se han reportado cambios de uso del suelo suscitados por procesos de deforestación, transición forestal, expansión de asentamientos y usos turísticos, reportándose la pérdida de bosques tropicales y el incremento de los usos agropecuarios (Nájera et al., 2010; Nené-Preciado et al., 2017; Romero et al., 2020).
En los tramos SCA se observaron alteraciones en las orillas del río relacionado con la presencia de bordos para evitar la inundación, los cuales impiden que la llanura de inundación no reciba agua e intercambio de sedimento durante las inundaciones de la temporada de lluvia. La construcción de estas obras afecta la morfología del cauce, modifica el índice de sinuosidad, confina el agua y los sedimentos en el cauce, limitando la exportación de sedimentos hacia la planicie de inundación con lo que se favorece el arrastre de sedimentos hacia la cuenca baja (Gálvez-Marroquín, 2017). Esta modificación en el balance de sedimentos (tamaño y tipo de partículas) influye en la distribución de la vegetación a lo largo del río y modifica la estabilidad del corredor de vegetación ribereña, que es una condición esencial que protege al cauce de la erosión durante eventos de avenidas extraordinarias (Gordon y Meentemeyer, 2006), estas causas contribuyen a que en estos tramos los valores de RQI sean más bajos.
Los valores bajos del RQI en los tramos SCA, se relacionan además con una mayor discontinuidad longitudinal en la vegetación ribereña que los tramos S, debido al desarrollo de actividades agrícolas y ganaderas en los mismos. Las zonas agrícolas y con pastizales aledañas al río influyen negativamente en los valores del índice, ya que disminuyen el grado de conectividad de la zona de ribera con la vegetación natural adyacente, limitando el movimiento de la fauna silvestre y pudiendo provocar extinciones locales (González-del Tánago y García-de Jalón, 2006). También el puente existente (tipo de puente viga losa; Rodríguez-Hinostroza, 2019) en estos tramos (p.ej., SCA2 y SCA4) produce bajos valores del índice, ya que contribuye a generar procesos erosivos puntuales, modifica el grado de naturalidad del canal fluvial, actúa como rutas de invasión de especies exóticas y es fuente de contaminación química por el escurrimiento que genera (Fernández et al., 2009; Rodríguez-Téllez et al., 2016).
En este estudio, se encontró evidencia que indica que el tipo de cobertura adyacente a la ribera del río incide sobre los atributos de la vegetación como la riqueza, la composición de especies y la densidad de individuos. Consistente con Gantes et al. (2014), la vegetación ribereña fue vulnerable a los impactos antropogénicos relacionados con el cambio de uso del suelo, resultante de la expansión de las tierras con pastizales para alimentación del ganado.
En general, hubo evidencia de una fragmentación de la vegetación en los tramos SCA, los cuales presentaron menor ancho promedio de la vegetación ribereña en el corredor, así como menor número de individuos por hectárea lo que puede ser explicado por los tipos de suelo presentes en estos sitios (Aguiar y Ferreira, 2005; Méndez-Toribio et al., 2014). Una gran cantidad de información sugiere que el uso del suelo juega un papel importante en la explicación de la fragmentación y los patrones de cobertura ribereña (Corbacho et al., 2003; Méndez-Toribio et al., 2014). Esta situación constituye un punto de especial preocupación para la planificación y gestión de estos tramos, considerando que la fragmentación del hábitat tiene efectos negativos sobre la viabilidad de las semillas y plántulas, afectando directamente la reproducción vegetal y la estructura, la composición y el funcionamiento de los ecosistemas (Carrasco et al., 2014).
La mayor riqueza de especies se encontró en los tramos VSA y S, en comparación con los tramos SCA. Esto puede estar relacionado con los efectos del cambio en el uso del suelo debido al aumento de la perturbación humana (Díaz-Pascacio et al., 2018; Méndez-Toribio et al., 2014). Se ha reportado que a medida que aumenta la perturbación en un área, la comunidad de plantas exhibe una disminución en la calidad florística (Bowers y Boutin, 2008). Ferreira et al. (2005) reconocieron que la estructura leñosa riparia estaba relacionada con importantes alteraciones en el uso de la tierra y, evidenciaron que, tanto la proximidad como la extensión de los parches de uso del suelo, interactúan para influir en el grado de cambios en la estructura ribereña.
Los estimadores de diversidad indicaron, en todos los casos, que la riqueza potencial de especies en el área estudiada es más alta. En apoyo a los hallazgos de Méndez-Toribio et al. (2014), se sugiere que la riqueza puede limitarse a pequeños restos de vegetación a lo largo de la orilla del río. Por esta razón, el estudio en mayor detalle de la flora de este tipo de comunidad es esencial, especialmente dentro de los tramos mejor conservados (S y VSA). Aún debe completarse la descripción de la flora local e identificación de especies que tienen algún riesgo de desaparecer, así como identificar aquellas especies con potencial de uso para restaurar tramos degradados.
El número total de especies observadas (90) resultó ser baja en comparación con lo encontrado por Díaz-Pascacio et al. (2018) para el río Sabinal quienes registraron 143 especies en una cuenca con predominio de bosques tropicales bajos caducifolios y bosques tropicales subcaducifolios. Los resultados pudieran atribuirse a la discontinuidad en la vegetación de algunos de los tramos estudiados, o a la eliminación de la vegetación aledaña a las zonas riparias para establecer pastos para la alimentación del ganado (Camacho-Rico et al., 2006) lo que origina un menor grado de conservación en comparación con otras localidades. Sin embargo, también pudiera estar dado a que el diseño de muestreo empleado en este estudio, así como el tipo de vegetación y ambiente son diferentes al estudio de Díaz-Pascacio et al. (2018), donde se muestrearon todos los árboles de los tramos y en 3 subcuencas diferentes, lo que pudo determinar que encontraran mayor número de especies.
Por otro lado, el número de especies es alto en comparación con el río La Sauceda, Durango, México, donde se reportaron 21 especies (Rodríguez-Téllez et al., 2016) y con el río Tembembe en Morelos, México donde se reportaron 74 especies en una zona de transición entre un clima templado subhúmedo y otro cálido subhúmedo (Camacho-Rico et al., 2006). Por otra parte, es similar a lo encontrado por Lott et al. (1987), quienes identificaron 105 especies en Chamela, México, donde predomina un bosque tropical subcaducifolio y por Ortiz (2019) en el río Ayuquila-Armería, Jalisco, México, quien reportó 102 especies, donde predomina el bosque tropical caducifolio. No obstante, las comparaciones con estos estudios deben tomarse con reservas, pues los métodos empleados, los tipos de ambientes y de vegetación son diferentes a los de esta investigación.
Especialmente en los tramos SCA se registró la disminución de la densidad de individuos por hectárea lo que pudiera estar relacionado con la perturbación generada por actividades antrópicas desarrolladas en las proximidades de la orilla del río, así como el efecto de borde. Las actividades antrópicas incluyeron la tala selectiva de especies, la eliminación parcial de comunidades vegetales y el ganado alimentándose en las orillas del río. Estas actividades pueden afectar la dominancia relativa y la distribución de las especies, causando la extinción local de especies raras debido a su baja densidad y abundancia (Aguiar y Ferreira, 2005).
Además de las actividades antrópicas, el efecto de borde puede considerarse un factor ecológico de importancia que puede explicar parcialmente los resultados obtenidos (Murcia, 1995). Este efecto surge de la transición abrupta de un área con vegetación a pastizal o hacia un área urbana y provoca la modificación del microclima (p. ej., temperatura del aire, humedad del suelo, déficit de presión de vapor e intensidad de luz) en el continuo de vegetación (Saunders et al., 1999). Estos cambios de las condiciones ambientales provocan que los individuos estén al límite de su tolerancia fisiológica afectándose de esa forma su desempeño ecológico en términos de la supervivencia, crecimiento y reproducción (Violle et al., 2007). Sin embargo, las especies pioneras son favorecidas por estas condiciones, y aumentan su densidad cerca del borde (Rodríguez, 1997). Dadas estas condiciones, debe considerarse a las especies herbáceas y leñosas nativas, como precursoras en la recuperación de áreas degradadas, siendo las heliófitas de ciclo corto (pioneras) y las heliófitas perennes de crecimiento rápido, las más recomendables para ello (Mejía, 2005).
El Permanova mostró que la comunidad de plantas en los tramos S difiere de los tramos SCA. Esto puede explicarse por el cambio en el tipo de cobertura, las diferencias en factores de disturbio como la distancia a las zonas agropecuarias, variables topográficas como la altitud y la pendiente, y variables climáticas como la temperatura lo cual se evidenció con el NMDS (Fig. 5). Al respecto, Hough-Snee et al. (2015) demostraron que la composición de la vegetación ribereña se correlacionó fuertemente con los filtros ambientales a escala del paisaje, incluida la elevación, la precipitación y la temperatura. Un estudio previo realizado por Martínez-Camilo et al. (2018) en una zona cercana de la sierra Madre de Chiapas, en la vertiente del Pacífico indicó que la diversidad de árboles presenta un patrón de disminución de la riqueza conforme se incrementa la elevación, lo que se correlacionó con la temperatura, la lluvia y las tendencias de menor perturbación humana y que hay más especies en terrenos bajos, pero en su mayoría corresponde a especies pioneras, mientras que las tardías están en zonas más altas y con menos perturbación (Martínez-Camilo et al., 2018).
Otros autores han señalado que en las comunidades tropicales la diversidad y la estructura de la vegetación varían a través del gradiente altitudinal (Sánchez-Gutiérrez et al., 2021). Se ha demostrado que los gradientes altitudinales están asociados con cambios en las variables climáticas y en las características de la vegetación ribereña, incluyendo su diversidad, así como sus propiedades estructurales y funcionales (Gregory et al., 1991; Vannote et al., 1980; Ward et al., 2002). La vegetación ribereña cambia además, por el efecto de las actividades antrópicas que modifican el paisaje (Moreno-Jiménez et al., 2019), lo cual provoca cambios en la comunidad de especies vegetales. Esto puede explicar las diferencias observadas en la comunidad de plantas de los tramos S con los de SCA, pues los primeros se encuentran a mayor altura y alejados de zonas agropecuarias, caminos y poblados, lo cual, además de la influencia del tipo de cobertura adyacente a la ribera, pueden haber influido en la composición de especies. Esto coincide con otros estudios donde se ha demostrado que las actividades agropecuarias y la urbanización influyen en la estructura y diversidad de árboles ribereños (Dummel y Pinazo, 2013; Pennington et al., 2010).
Por otro lado, los cambios observados en la composición de especies pueden explicarse por el hecho de que en los sitios SCA algunos procesos fluviales están alterados p. ej. la inundación de las riberas y la deposición de sedimentos. Estos procesos influyen en las etapas de vida de las especies ribereñas como son la dispersión de propágulos, el establecimiento de plántulas, el crecimiento y la reproducción (Goebel et al., 2012).
La familia Fabaceae fue la más representativa. Esta familia ha sido reportada también en otros estudios de vegetación ribereña tropical en México, debido al mayor número de especies en las zonas de estudio y por su mayor distribución en los bosques tropicales (Díaz-Pascacio et al., 2018; Maldonado-Sánchez y Maldonado-Mares, 2010; Moreno-Jiménez et al., 2017; Ortiz, 2019; Vázquez-Negrín et al., 2011). Por otra parte, la presencia de especies incluidas en alguna categoría de amenaza como: Croton guatemalensis (protección especial), Licania arborea (amenazada) y Vatairea lundellii (peligro de extinción) destacan la importancia de esta cuenca para la diversidad biológica de la vegetación ribereña en México y urge la necesidad de su conservación. La conservación de la vegetación ribereña en la cuenca Margaritas debería de hacerse con una visión integrada e incluir educación ambiental tanto para la población local como para el personal técnico, garantizar los inventarios y seguimiento cuantitativos y cualitativos a largo plazo, enmarcar las actividades económicas como la ganadería bajo gestión sostenible, y la restauración de las zonas degradadas (González et al., 2017).
Según el RQI, las zonas degradadas abarcan aproximadamente 30% de las riberas que se encuentran en estado moderado y otro 23% en estado pobre (con una moderada a alta alteración en varios de los 7 atributos evaluados, i.e., la composición de especies y la regeneración natural). Estas zonas requieren de medidas de rehabilitación o restauración para mejorar las funciones riparias hidrológicas y ecológicas. En este sentido, Díaz et al. (2007), sugieren que desde el punto de vista biológico las prácticas de manejo deben centrarse en el mantenimiento de la función ecológica y la resiliencia, que depende principalmente de las especies o de los grupos funcionales estructuralmente más importantes. En este caso, en el río Margaritas se debe dar prioridad durante la restauración a especies como Andira inermis, Salix humboldtiana, Vachellia cornigera (Apéndice 2) que fueron las especies más importantes reportadas en el estudio. Además se debe trabajar para eliminar o reducir los impactos en la zona ribereña de actividades antrópicas como la construcción de infraestructuras (bordos, caminos, viviendas), la agricultura y la ganadería. Esto pudiera lograrse promoviendo otras opciones de uso de las zonas de ribera, contando con alternativas e instrumentos de compensación ambiental que promuevan la reducción de las principales presiones (Meli et al., 2017).
El bosque de ribera constituye un elemento clave para la evaluación del estado ecológico de los ríos y es imperativo el desarrollo de indicadores biológicos para un manejo sustentable de estas áreas (Granados-Sánchez et al., 2006; Kotwal et al., 2008). En este sentido, el uso del RQI ha mostrado ser una buena alternativa para evaluar la condición ecológica en los ríos mexicanos, pues además de ser una forma rápida y sencilla de evaluar el estado de conservación de las riberas, también ayuda a la definición de opciones de manejo (González-del Tánago y García-de Jalón, 2006, 2011). El índice ha demostrado proveer de elementos sólidos para el desarrollo de cualquier iniciativa de manejo en una cuenca hidrográfica, de igual forma se podría evaluar el impacto de las acciones de manejo una vez realizadas. Por lo anterior, recomendamos el uso del RQI en la evaluación de la condición ecológica de los ríos en México.

