




![Estructura genética y aptitud ambiental de poblaciones de pasto banderita [Bouteloua curtipendula (Michx.) Torr.] en Chihuahua, México](/img/es/next.gif)





Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versión On-line ISSN 2448-6698versión impresa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.13 no.3 Mérida jul./sep. 2022 Epub 22-Ago-2022
https://doi.org/10.22319/rmcp.v13i3.5878
Review
Effects of phytoestrogens on the reproductive physiology of productive species. Review
a Universidad Autónoma Metropolitana. Unidad Iztapalapa, Departamento de Biología de la Reproducción, Ciudad de México, México.
b Universidad Nacional Autónoma de México (UNAM). Facultad de Medicina Veterinaria y Zootecnia, Departamento de Fisiología y Farmacología, Ciudad de México, México.
c UNAM. Facultad de Medicina Veterinaria y Zootecnia, Departamento de Medicina Preventiva y Salud Pública, Ciudad de México, México.
d Universidad Autónoma Metropolitana. Unidad Xochimilco, Departamento de Producción Agrícola y Animal, Ciudad de México, México.
e UNAM. Facultad de Medicina Veterinaria y Zootecnia, Departamento de Genética y Bioestadística, Ciudad de México, México.
f UNAM. Facultad de Medicina Veterinaria y Zootecnia, Departamento de Reproducción, Ciudad de México, México.
g UNAM. Facultad de Química, Departamento de Biología, Ciudad de México, México.
Phytoestrogens (PEs) are chemical compounds from the secondary metabolism of some plants, they have a potential effect on the reproductive parameters of domestic animals, acting as agonists or antagonists of estrogen receptors. The objective of this review is to know the effects produced by a diet rich in PEs on the reproductive physiology of slaughter animals. A systematic review was carried out in two databases using keywords related to the effects produced by the intake of PEs in the diet on the reproduction of animals for slaughter, only controlled studies developed in vivo were considered. Contradictory results were found, on the one hand, the intake of a high content of polyphenolic compounds from different fodders, in the bovine female, was related to the decrease in fertility, presence of abortions and ovarian cysts; on the other hand, the intake of the high content of PEs induced an increase in the semen quality of the males of the species: cattle, sheep and leporids, so these effects can be attributed to the concentration, the type of PEs, sex, species and even the breed of the animal.
Key words Phytoestrogens; Slaughter animals; Reproductive effects; Coumestans; Isoflavones; Lignans
Los fitoestrógenos (FEs) son compuestos químicos provenientes del metabolismo secundario de algunos vegetales, tienen un efecto potencial sobre los parámetros reproductivos de los animales domésticos, al actuar como agonistas o antagonistas de los receptores estrogénicos. El objetivo de esta revisión es conocer los efectos que produce una dieta rica en FEs en la fisiología reproductiva de los animales de abasto. Se realizó una revisión sistemática en dos bases de datos mediante el uso de palabras clave relacionadas con los efectos que produce la ingestión de FEs en la dieta, sobre la reproducción de animales de abasto, únicamente se consideraron los estudios controlados desarrollados in vivo. Se encontraron resultados contradictorios, por un lado, la ingesta de un alto contenido de los compuestos polifenólicos, provenientes de distintos forrajes en la hembra bovina, estuvo relacionado con la disminución de la fertilidad, presencia de abortos y de quistes ováricos; por otro lado, la ingesta de alto contenido de FEs indujo un aumento en la calidad seminal de los machos de las especies: bovinos, ovinos y lepóridos; por lo que, dichos efectos se pueden atribuir a la concentración, al tipo de FEs, sexo, especie e incluso a la raza del animal.
Palabras clave Fitoestrógenos; Animales de abasto; Efectos reproductivos; Coumestanos; Isoflavonas; Lignanos
Introduction
In livestock activities, the selection of feed represents one of the main production costs and is one of the most technically complex aspects for the search for nutritional satisfaction in the different stages of animal production (breeding, rearing, gestation, lactation, weaning, growth and fattening), considering their physiological state, age and breed1-4.
Generally, the feed of slaughter animals (SAs) is based on grains of sorghum and corn, since they provide a rich source of energy, in addition to other feeds that provide protein, such as soybeans, canola, alfalfa and clover2-4. It is essential to highlight that it has been suggested that changes be made in the feed or forage source in the event of alterations in the reproduction of SAs4.
Plant-based feeds are considered a rich source of phytoestrogens (PEs), nonsteroidal polyphenolic compounds derived from plant metabolism with conformational structure similar to 17-β estradiol (E2)3,5. When ingested by animals, they can act as selective modulators of estrogen receptors (ERs) and act as endocrine disruptors in an agonistic or antagonistic manner, depending on the dose ingested5,6, thus interfering with the synthesis, secretion, transport and metabolism of reproductive hormones, during embryonic development and in adult life6-11.
Nearly 100 PEs have been recognized; these are categorized, according to their chemical structure, into four classes: isoflavonoids (genistein, daidzein, formononetin); flavonoids (naringenin, kaempferol); coumestans (coumestrol “COU”), sativol, COU diacetate, 4-methoxycoumestrol) and lignans (enterolactone and enterodiol)12,13. Soybeans are the most abundant source of isoflavones, as it is one of the feeds with the highest content of genistein and daidzein, while alfalfa and clover contain a high amount of coumarins. It is worth mentioning that PEs are naturally present in plants, as glycosides, which are hydrolyzed to aglycone (which is the active form) catalyzed by enzymes present in the digestive system14,15.
Through several studies, it has been shown that the intake of PEs causes reproductive alterations in animals of both sexes, as well as temporary infertility syndromes16,17. However, there is also evidence that PEs favor reproduction because they increase the concentration, motility and volume of spermatozoa necessary for fertilization18-20.
Due to the economic and productive importance that reproductive alterations represent in the context of animal production, the objective of this work is to know the potential effect that feeds rich in PEs have on the reproduction of SAs.
Method
This document was developed in accordance with the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses)21 statement.
Literature search
A systematic review was conducted on the Web (Figure 1); Google Scholar and PubMed were used as specialized information search engines with the aim of identifying studies that explored the effects of PEs present in the diet on the reproductive physiology of SAs, which have been published in blind peer-reviewed journals, extended abstracts in specialty congresses, and in graduate theses between 1996 and 2019. The keywords were established according to the PICO (Participants, Interventions, Comparisons and Outcomes) principle: P as slaughter animals (cattle, sheep, goats, equines, pigs, leporids and poultry); I as feeding with a diet rich in PEs; C as control groups, or groups of animals with low phytoestrogen diet, or reproductive variables of the same experimental group reported before or after exposure to the high-phytoestrogen diet; and R as reproductive variables in males and females. The statement of the topic of the review question was developed as: Do phytoestrogens present in the diet produce adverse effects on reproductive variables in SAs? According to the keywords, Google Scholar was first searched and, subsequently, PubMed, dated until June 2020. Generally, the following words and Boolean operators were used, in order to identify the studies available on the web: {‘Reproduction’ AND ‘Animals’ AND ‘Phytoestrogens’ AND/OR ‘Fertility’ AND/OR ‘Feed’ AND/OR ‘Isoflavones’ AND/OR ‘Coumestans’, AND/OR ‘Lingans’ AND ‘Cows’ OR ‘Heifers’ OR ‘Bulls’ OR ‘Ewes’ OR ‘Small Rumninant’ OR ‘Sheep’ OR ‘Ovine’ OR ‘Goat’ OR ‘Nanny-goat’ OR ‘Equine’ OR ‘Mare’ OR ‘Horses’ OR ‘Porcine’ OR ‘Sows’ OR ‘Pigs’ OR ‘Leporids’ OR ‘Rabbits’ OR ‘Hens’ OR ‘Rooster’ OR ‘Chickens’} {‘Reproducción’, AND ‘Animales’, AND ‘Fitoestrógenos’ AND/OR ‘Fertilidad’ AND/OR ‘Alimentación’ AND/OR ‘Isoflavonas’ AND/OR ‘Coumestanos’ AND/OR ‘Lignanos’ AND ‘Vacas’ OR ‘Vaquillas’ OR ‘Toros’ OR ‘Pequeños rumiantes’ OR ‘Ovejas’ OR ‘Ovinos’ OR ‘Cabras’ OR ‘Equinos’ OR ‘Yeguas’ OR ‘Caballos’ OR ‘Porcinos’ OR ‘Cerdas’ OR ‘Lepóridos’ OR ‘Conejos’ OR ‘Aves’ OR ‘Gallinas’ OR ‘Gallos’ OR ‘Pollos’}. All the articles were searched in English and Spanish, were initially examined by reading their abstracts. The full texts of the documents included preliminarily were reviewed again to select the material to be used. Figure 1 shows a flowchart that details the selection of studies.
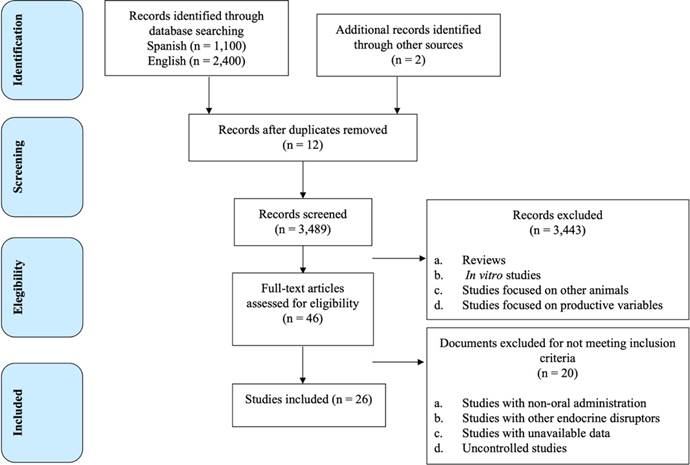
Inclusion and exclusion criteria
The studies considered were those on controlled experiments in vivo, in SAs fed a diet rich or supplemented with PEs (soybeans, clover, alfalfa) for more than seven days, and that included the determination of the amount of PEs ingested or sufficient information to calculate it, in that case, the amount of PEs in the diet (mg/kg of DM [dry matter]) was obtained from the database of the United States Department of Agriculture22,23; likewise, to obtain the amount of daily feed, the ratio of the animals’ weight and the amount of feed consumed containing PEs (reported in the article) was obtained. Those studies in which the administration of PEs was not related to feeding, or was administered orally through an excipient, or when reproductive parameters were not assessed, were excluded.
Data extracted from eligible studies were recorded as follows: species, breed, daily diet, main effects on reproductive variables, exposure time in days, PEs present in the diet and amount of PEs (mg/kg of DM).
Risk of bias analysis
The included papers were evaluated by two examiners in order to clarify emerging doubts, a third party was consulted to identify the presence of bias of randomization, blinding, results reporting and others (Table 1). To represent the bias analysis, a color scale was proposed: if the study met the described criterion, the green color was used; if the study was unclear or there was not enough information for the evaluation of bias, the yellow color was used.
Table 1 Summary of bias analysis of each included study
| Reference | The random selection of the animal sample is clearly described |
The researchers who selected the sample did NOT know to which treatment it would be assigned |
Blinding of evaluators to the assigned treatment was ensured |
The results are complete |
The study is free of other sources of bias |
|---|---|---|---|---|---|
| García et al., 201824 | |||||
| Woclawek-Potocka et al., 200525 | |||||
| Hashem et al., 201626 | |||||
| Rodríguez et al., 201327 | |||||
| Piotrowska et al., 200628 | |||||
| Yurrita et al., 201729 | |||||
| Cantero et al., 199630 | |||||
| Pace et al., 200631 | |||||
| Hashem et al., 201832 | |||||
| Pace et al., 201133 | |||||
| Aragadvay-Yungán et al., 20182 | |||||
| Sierra et al., 201534 | |||||
| Domínguez et al., 201419 | |||||
| Ferreira-Dias et al., 201335 | |||||
| Gentao et al., 199936 | |||||
| Yuan et al., 201237 | |||||
| Cardoso et al., 200738 | |||||
| Cardoso et al., 200939 | |||||
| Yousef et al., 200418 | |||||
| Hashem et al., 200840 | |||||
| Saleh et al., 201941 | |||||
| Ni et al., 200742 | |||||
| Lu et al., 201743 | |||||
| Wistedt et al., 201244 | |||||
| Arija et al., 200645 | |||||
| Heng et al., 201746 |
The green color indicates low risk of bias, the yellow color indicates that it is not clear or there is not enough information for the evaluation of the criterion, according to the authors’ judgment.
Results
In the systematic search, 26 specialized documents were finally collected: six on cattle (one article in Spanish and three in English, a master’s thesis in Spanish and a doctoral thesis in English); seven on sheep (one in Spanish and four in English, a master’s thesis in Spanish, a conference abstract in Spanish); one on equines in English; two on pigs in English; four on leporids in English and six on poultry in English (Table 2). All articles were published between 1996 and 2019.
Table 2 Summary of the main effects of phytoestrogens (PEs) on productive species
| Species | Condition and breed |
Daily intake | Main effects | Exposure (days) |
Amount of PEs according to the species and presentation of the plant (mg/kg of DM) |
Authors and year |
|---|---|---|---|---|---|---|
| Cattle | Holstein/Polish cows | 2.5 kg of soybeans | Decrease in PR | 300 | 1900 of GEN and DAI | Woclawek-Potocka et al., 200525 |
| Holstein cows | ~5 kg of alfalfa | Increase in E2, decrease in P4 and LH | 60 | ~25-65 of COU | Rodríguez et al., 201327 | |
| Holstein cows | ~5 kg of red clover | Increase in E2, decrease in P4 and LH | 60 | ~100 of GEN and 110 of DAI | Rodríguez et al., 201327 | |
| Holstein/Polish cows | 2.5 kg of soybeans | Decrease in P4 | 21 | 1900 of GEN and DAI | Piotrowska et al., 200628 | |
| Bradford heifers | 2.22 of kg soybeans | Decrease in PR, increase in abortions, not significant | 300 | ~570 of GEN and 310 of DAI | García et al., 201824 | |
| Holstein/Polish heifers | 2.5 kg of soybeans | No difference in PR | 21 | 1900 of GEN and DAI | Woclawek-Potocka et al., 200525 | |
| Holstein heifers | ~7 kg of clover | Decrease in PR and P4 Increase in E2 | 150 | 6.60 of GEN, 8.05 of DAI, 2.85 of FOR and 282.5 of BI | Hashem et al., 201626 | |
| Angus male calves | ~800 g of soybean meal | Increase in sperm concentration | 360 | ~135 of GEN and 6.32 of DAI | Yurrita et al., 201729 | |
| Sheep | Manchega ewes | Alfalfa pasture ad libitum | Endometrial and paraovarian cysts | 300 | 17-30 of COU | Cantero et al., 199630 |
| Rahmani ewes | 800 g of subterranean clover | Shorter estrus duration | 180 | 6.60 of GEN, 8.05 of DAI, 2.85 of FOR and 265 of BI | Hashem et al., 201832 | |
| Comisana female lambs | Subterranean clover pasture ad libitum | No effects on reproduction | 60 | 797 of GEN, DAI, FOR and BI | Pace et al., 200631 | |
| Sarda female lambs | Alfalfa pasture ad libitum | No alterations in reproductive system, fertility and fecundity | 60 | 900-10210 of GEN, BI and FOR | Pace et al., 201133 | |
| Sarda female lambs | Subterranean clover pasture ad libitum | No alterations in reproductive system, fertility and fecundity | 60 | 900-10210 of GEN, BI and FOR | Pace et al., 201133 | |
| Comisana female lambs | Subterranean clover pasture ad libitum | No alterations in reproductive parameters | 600 | 810-880 of GEN, BI and FOR | Pace et al., 201133 | |
| Creole rams | ~1 kg of alfalfa | Sperm concentration inverse to the dose | 45 | ~25-65 of COU | Aragadvay-Yungán et al., 20182 | |
| Hampshire/Suffolk rams | 1 kg of alfalfa meal | No alterations in sperm parameters | 90 | ~25-65 of COU | Sierra et al., 201534 | |
| Hampshire/Suffolk rams | 200 g of extruded soybeans | No alterations in sperm parameters | 90 | ~570 of GEN and 310 of DAI | Sierra et al., 201534 | |
| Katahdin/Pelibuey male lambs | ~230 g of alfalfa | No alterations in sperm parameters or morphology | 90 | ~25-65 of COU | Domínguez et al., 201419 | |
| Comisana male lambs | Subterranean clover pasture ad libitum | No effects on reproduction | 60 | 797 of GEN, DAI, FOR and BI | Pace et al., 200631 | |
| Equines | Lusitano mares | 1 kg of alfalfa pellets | High concentrations of COU in serum | 14 | ~25-65 of COU | Ferreira-Dias et al., 201335 |
| Pigs | Large White/Erhualian pregnant sows | ~1.5 mg of DAI | Increase in milk production | 37 | - | Gentao et al., 199936 |
| Minipig boars | ~6.75-27 mg of soybean isoflavones | At low doses, increase in testicular index; at high doses, decrease in testicular index, LH and T4 | 60 | 125-500 of ISO | Yuan et al., 201237 | |
| Leporids | New Zealand pregnant does | ~20 g of soybean meal | No effect on reproductive organs, semen quality and sexual behavior of F1 | ~256 | ~135 of GEN and 632 of DAI | Cardoso et al., 200738 |
| New Zealand bucks | ~32 g of soybean meal | No alterations in reproductive organs | 60 | ~135 of GEN and 632 of DAI | Cardoso et al., 200939 | |
| New Zealand bucks | 7.13-14.25 mg of ISO* | Improvement in the characteristics of semen and libido | 91 | - | Yousef et al., 200418 | |
| Line V bucks | ~15.44 g of soybeans | It does not affect semen fertility, decrease in T4 | 84 | 240.4 of DAI and 131 of GEN | Hashem et al., 201840 | |
| Line V bucks | ~19.3 g of flaxseed | It does not affect semen fertility, decrease in T4 | 84 | 368 of SECOI and 52.8 of DAI | Hashem et al., 201840 | |
| Poultry | Bovans Brown hens | ~120 mg of flax | Increase in LH, FSH, E2 | 42 | - | Saleh et al., 201941 |
| Bovans Brown hens | ~131 mg of fenugreek | Increase in LH, E2 | 42 | ~0.1 of DAI and 0.1 of GEN | Saleh et al., 201941 | |
| Bovans Brown hens | ~128 mg of flax and 128 mg of fenugreek | Increase in LH, FSH, E2 | 42 | ~0.1 of DAI and 0.1 of GEN | Saleh et al., 201941 | |
| ISA hens | ~1.2 mg of DAI | Increase in oviduct weight | 63 | - | Ni et al., 200742 | |
| Rugao hens | ~5.4-22.57 mg of DAI | No effect on fertility | 84 | - | Lu et al., 201743 | |
| Lohmann Brown hens | ~5 mg of DAI | No changes in morphology and size of ovaries and oviducts high sensitivity to DAI | 84 | - | Wistedt et al., 201244 | |
| Selected Leghorn hens | ~5 mg of DAI | No changes in morphology and size of ovaries and oviducts | 84 | - | Wistedt et al., 201244 | |
| Young breeders | ~0.75 mg of ISO | Increase in testicle weight, GnRH | 63 | - | Heng et al., 201746 | |
| Cobb chickens | ~2.38-7.14 mg of extruded beans | Increase in T4 and androstenedione | 21 | - | Arija et al., 200645 | |
| ~2.38-7.14 mg of raw beans | Decrease in T4 and androstenedione | 21 | - | Arija et al., 200645 |
BI= Biochanin A; COU= Coumestrol; DAI= Daidzein; E2= 17-ß estradiol; FOR= Formononetin; F1= Filial generation 1; GEN= Genistein; GLY= Glycitein; GnRH= Gonadotropin-releasing hormone; ISO= Isoflavones; LH= Luteinizing hormone; MCOU= Methoxycoumestrol; PEP= Para-ethyl-phenol; P4= Progesterone; SECOI= Secoisolariciresinol; T4= Testosterone; PR= Pregnancy rate; -= Not applicable; *= Every 3 days; ~= approximate calculation.
Cattle
In a study conducted on 11-month-old Bradford heifers (n=15 per group) fed, six months before their first service, with 0.8 % of soybeans, calculated based on live weight (LW), (feed that was considered by the authors to be high in PEs when compared to the usual feeding of 0.3 % LW), a percentage of gestation (%G) of 93 % and an abortion were observed, compared with the control group, where a 100 % success rate in pregnancy and no cases of abortion were reported; the authors mention that it is not possible to affirm that feeding with 0.8 % LW was the cause of abortion, and that this feeding did not affect reproductive parameters24. In another study, was evaluated the %G in Holstein/Polish heifers (75 and 25 %) that were constantly fed 2.5 kg of soybeans, it was observed that the pregnancy rate was not significantly different compared to the control group, although equol and p-ethyl-phenol (compounds derived from the metabolism of PEs) were detected in the serum of the heifers25. On the other hand, it has been reported that, in Holstein heifers fed clover for five months, the %G decreased (61.5 %), compared to heifers fed corn silage (92.3 %); the percentage of heifers that did not become pregnant after several inseminations was higher (38.46 %) in the group of cows fed clover compared to its control (7.7 %)26 (Table 2).
In adult Holstein/Polish cows (75 and 25 %) that were fed 2.5 kg of soybeans from a lactation period prior to mating, they had a %G of 60 % compared to the control group, which showed 100 % gestation25. In the lactation stage of Holstein cows fed for 60 d with alfalfa or red clover meal, alterations in the concentrations of E2, progesterone (P4) and luteinizing hormone (LH) were reported. A significant decrease in E2 was documented in the groups of cows fed alfalfa (2.32 ± 0.12 pg/mL) or red clover (2.25 ± 0.67 pg/mL) compared to control cows (4.24 ± 0.31 pg/mL); the concentration of P4 decreased at the end of the supplementation period in cows fed alfalfa (1.586 ± 0.27 ng/mL) or red clover (0.988 ± 0.3 ng/mL) compared to control animals (2.82 ± 0.34 ng/mL); LH also decreased in cows in the groups fed alfalfa (3.82 ± 0.22 IU/mL) or red clover (3.7 ± 0.26 IU/mL) compared to control cows (6.66 ± 0.39 IU/mL)27. In another study, it was observed that the concentrations of P4 decreased throughout the estrous cycle in Holstein/Polish black cows when they were fed 2.5 kg of soybeans for 21 d, compared to the control group, this suggests that PEs contained in soybeans can alter the function of the corpus luteum (CL), it is worth mentioning that the effects began to be observed between d 15 and 18 after its intake28 (Table 2). In the only available study conducted on Angus bulls, with a diet with 10 % of soybeans from weaning to pre-puberty, an improvement in scrotal growth and semen quality in adulthood was observed29.
The documented differences between sexes, derived from the effects produced by PEs, are biologically plausible, given that PEs are known xenoestrogens that alter the endocrine system depending on the availability of ERs and target organs of both sexes. In the studies reviewed, the age of exposure to PEs and the type of PEs determine their biological effect, observing a greater harmful effect in adult cows kept with feed rich in PEs than in heifers.
Small ruminants
In Manchega ewes that consumed alfalfa ad libitum for 10 mo and that contained COU at the rate of 25 ppm in autumn, 30 ppm in winter and 17 ppm in spring, it was found that 43 % of them showed alterations in the genital tract: cysts or microcysts in the endometrium accompanied by petechiae and ecchymosis in the uterine mucosa, increased glandular activity and paraovarian cysts30. There are studies in which no harmful effects on the reproductive parameters of ewes and males of the Comisana breed are documented, it has even been suggested that prolonged administration of subterranean clover, with low formononetin content (less than 10 % of the total isoflavones on a dry basis), induces a significant improvement in the weight gain of animals and, in males, good carcass and meat characteristics31. In another study conducted on pregnant Rahmani ewes, which were fed for two months prior to calving and until the induction of the next heat (3.5 mo postpartum) with 849.4 g/kg of DM of Trifolium alexandrinum, no significant differences between groups in ovarian activity and ovulation in induced heat were documented, but females fed Trifolium alexandrinum showed shorter estrus duration (20 h) compared to ewes fed corn (34 h), in which P4 concentrations in the luteal phase of the induced estrus were significantly higher, compared to treated ewes32.
In another work conducted on female lambs of the Sarda and Comisana breeds, the effects of feeding ad libitum with alfalfa or subterranean clover were evaluated, in these animals, no alterations were observed in the development of the reproductive system, fertility, fecundity, reproductive performance and calving interval, although animals fed clover had a higher weight at puberty, the authors suggest that some clover varieties do not negatively affect the reproduction of ewes and appear to improve the growth rate of animals33.
In Creole rams fed 1.1 kg/d of alfalfa contaminated with Pseudopeziza medicaginis at 10, 40 and 70 % for 45 d, a significant decrease in sperm concentration was documented in the rams fed alfalfa contaminated with the fungus at 40 % and 70 %, compared to the group with 10 %2. It should be mentioned that fungal infestations caused by Pseudopeziza medicaginis increase the synthesis of phytoestrogenic substances such as coumarins and isoflavones2. Other studies do not report differences between sperm characteristics, for example, in a study with hybrid Hampshire/Suffolk sheep, which were fed about 1 kg of alfalfa (2.5-6.5 mg of COU/100 g of alfalfa22,23) or 200 g of extruded soybeans (57 mg of genistein/100 g of extruded soybeans; 31 mg of daidzein/100 g of extruded soybeans22,23), daily for 90 d, no alterations were reported in sperm volume, color, motility and concentration, evaluated in fresh or cryopreserved semen34. Likewise, in three-month-old hybrid Katahdin/Pelibuey lambs fed 23 % of alfalfa for 90 d, no differences were observed in sperm volume, sperm membrane integrity, total or progressive motility and acrosome status19 (Table 2). In sheep, contradictions were found regarding the deleterious or beneficial effects attributed to the intake of PEs on reproductive variables.
Equines
The effects of PEs due to feeding on equines are poorly described. Conjugate and free (active) forms of COU and its metabolites have been identified in the plasma of mares fed for 14 d with alfalfa pellets in increasing concentrations of up to 1 kg/d, during the time of the experiment, all mares cycled, on day zero half of the mares were in the follicular phase, and the other half in the luteal phase, on d 13 and 14 of the experiment, all the mares had the luteal phase35 (Table 2). The results suggest that PEs affect the length of the estrous cycle and can prolong luteal function in the mare, due to the induction of the persistence of CL35.
Pigs
In pregnant sows fed a regular diet supplemented with 0.005 mg of daidzein/kg of LW, in the peripartum period (from 30 ds prepartum to d 7 postpartum), they showed an increase in the weight of the litter compared to the control group, which did not consume daidzein; in addition, the production of milk, colostrum proteins and growth hormone increased by about 12 % in those sows fed that isoflavone36. In minipigs fed low concentration of isoflavones (250 ppm), no negative effects on reproduction were observed; in contrast, when the concentration increased (500 ppm), the testicular index ((bilateral testicular weight/total body weight) x 100 %) LH and testosterone (T4) decreased; in addition, there was an increase in apoptotic germ cells, indicating testicular peroxidation37 (Table 2).
In sows, feeding supplemented with daidzein favors the weight of the offspring at calving and milk production, in the case of males, the effect of PEs in the diet depends on the amount administered, low amounts of PEs favor testicular function.
Leporids
In a study conducted on pregnant does fed a diet of 18 % of soybean meal (13 mg/kg of isoflavones/body weight) during the stages of gestation, lactation and until the offspring were 33 weeks old, it was observed that males were earlier in the time of onset of puberty compared to the control group, no significant differences were reported in the morphology of the reproductive organs, semen quality and sexual behavior38. In another study, the effect of exposure to a commercial rabbit diet with 18 % of soybean meal during perinatal period (intrauterine and lactation) on the morphology of the reproductive organs of males at 26 wk of age was evaluated. No alterations were observed in the reproductive tract of the male progeny39.
In 7-mo-old adult New Zealand male rabbits fed 30 % of Berseem (Trifolium alexandrinum) hay and supplemented with 2.5-5 mg of isoflavones/kg of body weight every third day for 13 wk, improvements in semen and libido characteristics were documented, since there was an increase in sperm volume, concentration and percentage of sperm motility18. When the effect of a diet rich in soybeans (80 g of soybeans/kg of feed) and flaxseed (100 g of flaxseed/kg of feed) in adult rabbits was evaluated, increases in the occurrence of abnormalities in spermatozoa, decrease in libido and in the process of steroidogenesis were observed, however, when they were crossed with untreated females, pregnancy rate, litter size and fertility were not affected40 (Table 2).
The results of the previous studies suggest that the administration of commercial feed, with a concentration of up to 18 % of soybeans, to pregnant does, does not produce, in the offspring, alterations in the morphology of the reproductive organs in F1 males. Nor do diets with percentages less than 80 g of soybean meal or 100 g of flaxseed/kg of feed cause effects on the reproductive organs of males, the volume of the ejaculate or fertility. No studies evaluating the effects of PEs directly on the reproductive physiology of females were found (Table 2).
Poultry
In a study conducted on 65-wk-old Bovans Brown laying hens41 fed diets supplemented with 1 g of flax seeds or fenugreek/kg of feed (rich in PEs, 0.20 mg of isoflavones/kg of DM) for 6 wk, an increase in the concentrations of E2 and LH, weight and an improvement in shell thickness at the end of the egg laying cycle were observed, both with the supplementation of the seeds separately and with their combination (flax and fenugreek)41. The increase in eggshell thickness documented in the previous study was also present in another, in which 445-d-old ISA laying hens were fed a diet supplemented with 10 mg of daidzein/kg of feed42. In addition, in this last study, a significant increase in the proportion of oviduct weight to body weight was reported42. There were no alterations in the width and length of the egg, nor in the serum concentrations of E242. On the other hand, it has been observed that in 44-wk-old Rugao laying hens fed supplemented diets from 60 to 248 mg of daidzein/kg of feed for 12 wk, it did not generate significant differences in egg quality or fertility, although an increase in the hatchability of egg laying was observed43. Wistedt et al44 also reported the absence of effects on reproductive variables in 15-wk-old Lohmann Selected Leghorn (LSL) and Lohmann Brown (LB) laying hens on the morphology and size of ovaries and oviducts after being supplemented with 50 mg of daidzein/kg of feed. A difference in the sensitivity of the breeds to daidzein was observed, since the eggshells were thicker in LB hens than in LSL hens44. In 1-d-old Cobb broilers, fed 100, 200 or 300 mg of extruded or raw beans/kg of feed for 21 d, a decrease in T4 and androstenedione concentrations was observed with raw bean feeding, in contrast, the administration of extruded beans increased the same variables45.
Finally, in 70-d-old young breeding roosters fed a commercial product added with isoflavones, it was observed that, with 5 mg/kg, testicle weight, concentration of gonadotropin-releasing hormone (GnRH) and expression of the mRNA of the StAR enzyme46 were increased (Table 2). The results of the studies suggest that PEs cause favorable effects on reproductive variables of male and female poultry.
Bias
In none of the studies included in the present review, the evaluators were blinded to the assigned treatment, nor in the selection of the sample. In contrast, complete results were identified in all the papers, and they were free from other sources of bias. More than half of the included studies used some randomization technique for their experimental units18,27,30,31,36,37,38,39,40,41,42,43,45,46.
Discussion
In ruminants, isoflavones are metabolized in the rumen, which generates estrogenic or non-estrogenic compounds. Secoisolariciresinol and matairesinol are the precursors of COU, enterodiol and enterolactone; biochanin A and genistein can be broken down into p-ethyl-phenol (non-estrogenic); formononetin is metabolized to daidzein and finally to equol, a more estrogenic compound that is absorbed through the rumen wall47,48.
The main effects produced by PEs in cattle include hormonal alterations, decrease in %G and increase in abortions, this depends on the reproductive stage, the time of consumption and the type of PEs involved. Early embryonic mortality and the increase in the abortion rate can be explained by the ability of PEs to inhibit the secretion of P4 stimulated by LH49,50. In cows, the release of prostaglandins (PG) with luteolytic action is regulated by E2 and P451. PEs and their active metabolites alter the PGE2-F2α ratio, which leads to the non-physiological production of luteolytic agents during gestation and the estrous cycle25. In gestation, the balance between PGE2-F2α is crucial for the maintenance and function of CL, the recognition of pregnancy, embryo implantation and development, so the stimulation of PGF2α production can lead to interfere with this balance and embryonic development52. PEs as agonists, in non-pregnant animals, reduce the length of the estrous cycle, since during luteolysis and ovulation, stimulation of PGF2α secretion can accelerate positive feedback between PGF2α and oxytocin51,52. On the other hand, PEs, as antagonists, induce alterations in follicular development and therefore, the absence of estrus53,54. In addition, a possible positive relationship between the concentration of isoflavones in blood plasma and the incidence of silent heat in dairy cattle fed soybeans has been reported55.
Being structurally similar to E2, PEs act as agonists or antagonists of ERs7,56. The biological effects attributed to PEs occur with concentrations around 1,000 times higher than endogenous E2 concentrations (1-10 nM). This is based on the fact that Woclawek-Potocka et al reported 1.6 ± 0.3 μM of p-ethyl phenol and 1.2 ± 0.28 μM of equol25; Piotrowska et al reported 1.28 ± 0.10 μM of equol and 6.24 ± 0.30 μM of p-ethyl phenol28; Zdunczyk et al reported daidzein, genistein, equol and p-ethyl-phenol in a range of 0.1-3.6 μmol/L in cows that showed reproductive alterations55. In another study, it was calculated that the intake of 66.8 mg/kg of COU (present in alfalfa) produced plasma concentrations of 13 ng/mL of this compound, which also turns out to be 1,000 times higher than the concentration of E2 during estrus25. Even considering that the biological activity of COU is 160 times lower compared to that of E2, the amount would be equivalent to six times the effective concentration of E2 in estrus, and therefore, sufficient to induce estrogenic changes similar to those found in the cow during this phase of the estrous cycle57. The above data suggest that cows that are continuously exposed to a diet that includes PEs may show reproductive alterations, contrary to what is reported in males, in which feeding with 10 % of soybeans as a source of protein improves sperm formation and concentration29.
In sheep, the effects of PEs on reproductive variables are contradictory. In some studies, it has been suggested that these compounds do not cause alterations as in other species19,20; in others, their deleterious effect is evident, even the “Clover Syndrome” has been described, which consists of infertility, prolapse of the uterus and dystocia58,59. The main effects reported in females are morphological alterations in reproductive organs and increased activity of the endometrial glands, which leads to quantitative and qualitative changes in cervical mucus, which can hinder fertilization30,60. It is reported that PEs sensitize the cervix to estrogenic action, in addition to occupying ERs in the cervix, they also stimulate the appearance of new binding sites61. In addition, PEs alter the secretion of the follicle-stimulating hormone (FSH)62. In ewes, the number of recruitable follicles depends on FSH concentrations63, therefore, PEs are likely to interfere with follicular recruitment.
It has been reported that the factors related to the absence of effects caused by PEs in some studies are seasonality, the dose of PE ingested and the species and conditions of the plant used, since it has been documented that, in a single plant species, more than one type of PEs can be found at different concentrations; for example; under normal conditions, the concentrations of COU in alfalfa are 1 to 2 mg/kg and in circumstances of defense of the plant, the concentration of that PE can increase up to 100 mg/kg64. The concentration of PEs also depends on the presentation and organ of the plant, for example: the isoflavone content in raw green soybeans is 48.9 mg/100 g of DM; that of extruded soybeans is 91 mg/100 g of DM and textured soybeans contain 172.6 mg/100 g of DM22,23. With respect to flavonoids, mature soybeans contain 37.41 mg/100 g of DM and green soybeans contain 1.23 mg/100 g of DM22,23.
The transition of photoperiods contributes to the regulation of estrous behavior, ovulation, ERs availability and concentration of endogenous steroids32. In sheep, it has been suggested that prolonged ingestion of subterranean clover with PE levels below 0.3 % or ~10 mg/g DM of alfalfa does not produce infertility or reproductive disorders, it even significantly increases body growth33, probably because PEs also stimulate growth hormone42,65. It has been suggested that the differences in deleterious effects on reproduction may be due to the type of isoflavones administered, the difference in the number of ERs and the type of metabolism between the different species66. In male sheep, contradictory results were also found, Aragadvay et al2 describe reproductive alterations with the feeding of alfalfa contaminated with Pseudopeziza medicaginis at 40 and 70 %; other studies do not report harmful effects19,34. The reason why this difference in results exists is not described yet, probably due to the difference in the sensitivity of steroidogenic compounds between breeds of the same species, as reported in other animal models67, it is even likely that male sheep are less susceptible to the effect of phytoestrogens20 or that some plant varieties do not negatively affect the reproduction of ewes20,33.
According to the criteria established in this review, no published studies related to goats were found. However, it has been reported that goats constantly fed 30 % of dehydrated alfalfa had an incidence of 20 % of rectal or vaginal prolapses in the last month of gestation68.
In a case report of mares fed 5-8 kg/d of mixture of alfalfa and clover hay for at least 5 mo, uterine edema, absence of ovulation and accumulation of uterine fluid and return to normal ovarian cyclicity within 2 to 3 wk after withdrawal of the feed rich in PEs, were observed35. The mechanism of absorption of PEs is not fully described; in this species, digestion is very fast and feed can pass through the stomach and small intestine within the first 5 h69. This was confirmed in a study in which active forms of COU reached their highest level between 1 and 3.5 h after ingestion35. The amount of PEs in the pasture depends on the season of the year, since a decrease in COU and its metabolites was found when evaluated from November to March and it is known that its synthesis increases under adverse conditions for plants35. It is worth mentioning that the dehydration of alfalfa to produce pellets could reduce estrogenic activity70. Therefore, the exposure time, the presentation of PEs and the season of the year are fundamental for the effects to occur.
The effects of PEs, present in the diet of pigs, on reproductive variables are few, in contrast to the amount of data on estrogenic effects of feed contaminated with mycotoxins, or works in which the effect of isoflavones on meat quality and growth is studied71-73. Daidzein and genistein are the main isoflavones contained in soybean meal, which is the basic protein ingredient in the diet of pigs, and it is known that both represent 88 % of the isoflavones circulating in the blood73. One of the effects of feeding sows with diets containing PEs in the last third of gestation is an increase in the weight of animals, this may be due to the positive effect produced by PEs on growth hormone concentrations74. In males, the effect is much more evident, since, in the fetal stage, the concentrations of circulating estrogens are low, and the presence of PEs would stimulate growth due to a change in metabolism, proliferation and differentiation of skeletal muscle75. The increase in milk production and better milk quality in sows fed isoflavones could be due to their agonist effect on insulin-like growth factor-1 (IGF-1) and prolactin reported in periparturient sows76 or to a better balance of antioxidants77. In males, a positive effect on reproduction has been identified when low concentrations of PEs are administered37. This has been attributed to the binding of isoflavones to ERs in the hypothalamus, pituitary gland and testicles, thus stimulating spermatogenesis, sperm maturation and gonadal growth6.
As for leporids, it is reported that females are more sensitive to the effect of environmental estrogens38. In male rabbits, intrauterine and lactational administration of PEs do not induce deleterious effects on semen production or sexual behavior, it has even been suggested that soybean meal can be used as part of the normal diet in pregnant females without reproductive damage to the progeny38,39. The beneficial influence of isoflavones on semen parameters has been attributed to their antioxidant effect because they reduce the production of hydrogen peroxide and protect spermatozoa against oxidative damage18. In addition, in adult males, estrogen plays a major role in the prevention of the apoptosis in germ cells through its paracrine or autocrine actions in the testicles32. The type of PEs consumed by rabbits also influences the final effect, for example, lignans suppress T4 synthesis more strongly during spermatogenesis and libido, compared to isoflavones32.
In poultry, as in other domestic animals, PEs can act as agonists/antagonists, depending on the dose, tissue type, ER subtype and presence of endogenous hormones, therefore they are considered selective modulators of ERs78. In poultry, the effects produced by the consumption of PEs are related to the genetic component, since lines of LB and LSL hens showed different sensitivity to these compounds44; this could be due to the fact that there is a difference in the location and expression of ERs ß in uterine capillaries between genetic lines, which could influence the obtaining of greater weight of the oviduct44.
In bird males, it was observed that a diet rich in PEs promotes testicular growth due to the increase in hormonal secretion, probably due to the regulatory participation of isoflavones in the feedback mechanism of the hypothalamic-pituitary-gonadal axis79, which in turn regulates the secretion of FSH, LH and T4 that promotes the growth and maturation of Sertoli and Leydig cells. The administered dose also has an important role, it is documented that genistein has partial agonist effects in roosters at doses of 50-200 mmol/kg, and acts as an antagonist at doses of 400-500 mol/kg78; in addition, PEs have the ability to inhibit the activity of steroidogenic enzymes, and influence the viability of sex hormones through the regulation of their binding proteins; they alter brain centers related to sexual behavior as they cross the blood-brain barrier and bind to ER α and ß80.
Finally, the metabolism of dietary PEs is predominantly determined by gastrointestinal bacterial metabolism and depends on the reproductive stage, for example, during gestation, genistein has the potential to influence fetal metabolism and growth; in the colon, genistein can be metabolized to dihydrogenysteine or 6’-hydroxy-O-desmethylangolensine, while daidzein can be reduced to dihydrodaidzein and converted into O-desmethylangolensin or equol, these metabolites can be absorbed or metabolized into phenols in the colon lumen81. After the consumption of PEs, these are deconjugated by the intestinal bacterial flora, reabsorbed, re-conjugated in the liver and excreted in the urine25,61. The demethylation of these compounds occurs in the intestine by acetogenic bacteria and in the liver81. Therefore, the intestine microbiome present in each species will influence the final effect of PEs on productive and reproductive variables.
Conclusions
Based on the papers reviewed and discussed, it is concluded that phytoestrogens cause alterations in the reproductive physiology of slaughter animals considering four factors: 1) The plant species consumed; 2) The season of the year in which the plant species is consumed; 3) The particularities of the animals (metabolism, age, breed, species and sex) and 4) The processing conditions of the plant. Future studies should be carried out to elucidate the endocrine mechanisms of the actions of PEs in the reproduction of animals. It is necessary to reevaluate the ingredients that make up the feeds of the different productive animals.
Literatura citada
1. Meléndez P, Bartolomé J. Advances on nutrition and fertility in dairy cattle: Review. Rev Mex Cienc Pecu 2017;8(4):407-417. [ Links ]
2. Aragadvay-Yungán R, Novillo-Rueda M, Núñez-Torres O, Rosero-Peña Herrera M, Lozada-Salcedo E. Calidad seminal de carneros alimentados con dietas que contienen alfalfa (Medicago sativa) contaminada con Pseudopeziza medicaginis. Rev Ecua Invest Agrope 2018;2(1):14-19. [ Links ]
3. Pérez-Rivero JJ, Setién A, Martínez-Maya JJ, Pérez-Martínez M, Serrano H. Los fitoestrógenos y el efecto de su consumo en diferentes órganos y sistemas de animales domésticos. Agric Tec 2007;67(3):325-331. [ Links ]
4. Magnusson U. Environmental endocrine disruptors in farm animal reproduction: research and reality. Reprod Domest Anim 2012;47(Suppl 4):333-337. [ Links ]
5. Di Gioia F, Petropoulos SA. Phytoestrogens, phytosteroids and saponins in vegetables: Biosynthesis, functions, health effects and practical applications. Adv Food Nutr Res 2019;90:351-421. [ Links ]
6. Adams NR. Detection of the effects of phytoestrogens on sheep and cattle. J Anim Sci 1995;73(5):1509-1515. [ Links ]
7. Almstrup K, Fernández MF, Petersen JH, Olea N, Skakkebæk NE, Leffers H. Dual effects of phytoestrogens result in u-shaped dose-response curves. Environ Health Perspect 2002;110(8):743-748. [ Links ]
8. Cederroth CR, Zimmermann C, Nef S. Soy, phytoestrogens and their impact on reproductive health. Mol Cell Endocrinol 2012;355(2):192-200. [ Links ]
9. Ropero AB, Alonso-Magdalena P, Ripoll C, Fuentes E, Nadal A. Rapid endocrine disruption: environmental estrogen actions triggered outside the nucleus. J Steroid Biochem Mol Biol 2006;102(1-5):163-169. [ Links ]
10. Shanle EK, Xu W. Endocrine disrupting chemicals targeting estrogen receptor signaling: identification and mechanisms of action. Chem Res Toxicol 2011;24(1):6-19. [ Links ]
11. Whitten PL, Patisaul HB. Cross-species and interassay comparisons of phytoestrogen action. Environ Health Perspect 2001;109(Suppl 1):5-20. [ Links ]
12. Nilsson S, Makela S, Treuter E, Tujague M, Thomsen J, Andersson Gr, et al. Mechanisms of Estrogen Action. Phys Rev 2001;81(4):1535-1565. [ Links ]
13. Wocławek-Potocka I, Mannelli C, Boruszewska D, Kowalczyk-Zieba I, Waśniewski T, Skarżyński DJ. Diverse effects of phytoestrogens on the reproductive performance: cow as a model. Int J Endocrinol 2013;2013:650984. [ Links ]
14. Bonilla CA. Isoflavonas en ginecología, terapia no convencional. Rev Colomb de Obstet y Ginecol 2004;55(3):209-217. [ Links ]
15. Wang H, Murphy PA. Isoflavone Content in Commercial Soybean Foods. J Agric Food Chem 1994;42(8):1666-1673. [ Links ]
16. Peña-Corona S, León P, Mendieta E, Villanueva M, Salame A, Vargas D, et al. Effect of a single application of coumestrol and/or dimethyl sulfoxide, on sex hormone levels and vaginal cytology of anestrus bitches. Vet Méx 2019;6(1):1-15. [ Links ]
17. Pérez-Rivero JJ, Martínez-Maya JJ, Pérez-Martínez M, Aguilar-Setién Á, Serrano H. Efecto del coumestrol sobre la producción espermática y la conducta de exploración olfatoria de perros estimulados con moco vaginal estral. Vet Méx 2009;40(1):9-16. [ Links ]
18. Yousef MI, Esmail AM, Baghdadi HH. Effect of isoflavones on reproductive performance, testosterone levels, lipid peroxidation, and seminal plasma biochemistry of male rabbits. J Environ Sci Health, Part B 2004;39(5-6):819-833. [ Links ]
19. Domínguez-Rebolledo AE, Alcaraz-Romero A, Cantón-Castillo JG, Loeza-Concha H, Ramón-Ugalde J. Efecto de la Alfalfa (Medicago Sativa L.) en la dieta sobre la calidad de los espermatozoides epididimarios de ovinos Katahdin con Pelibuey. Reunión Científica Tecnológica, Forestal y Agropecuaria Tabasco 2014 y III Simposio internacional en producción agroalimentaria tropical. Tabasco: Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias 2014:186-189. [ Links ]
20. Valderrabano J. Alteraciones reproductivas asociadas al consumo de fitoestrógenos. Investigación Agraria Producción y Sanidad Animal 1992;7(2):115-124. [ Links ]
21. Moher D, Liberati A, Tetzlaff J, Altman DG, PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. PLoS Med 2009;6(7):e1000097. [ Links ]
22. Bhagwat S, Haytowitz DB, Holden JM. USDA database for the isoflavone content of selected foods, release 2.0. Maryland: US Depart of Agricult 2008;15. [ Links ]
23. Bhagwat S, Haytowitz DB, Holden JM. USDA database for the flavonoid content of selected foods, Release 3.1. US Depart of Agricult: Beltsville, MD, USA. 2014. [ Links ]
24. García DC, Martín AA, Vella MA, Nasca JA, Roldan Olarte EM. Evaluación de la alimentación con distintos niveles de inclusión de soja en la recría de vaquillonas. Vet Arg 2018;35(357)1-9. [ Links ]
25. Woclawek-Potocka I, Acosta TJ, Korzekwa A, Bah MM, Shibaya M, Okuda K, et al. Phytoestrogens Modulate Prostaglandin Production in Bovine Endometrium: Cell Type Specificity and Intracellular Mechanisms. Exp Biol Med 2005;230(5):326-333. [ Links ]
26. Hashem NM, El-Azrak KM, Sallam SMA. Hormonal concentrations and reproductive performance of Holstein heifers fed Trifolium alexandrinum as a phytoestrogenic roughage. Anim Reprod Sci 2016;170:121-127. [ Links ]
27. Rodríguez MCE. Determinación del contenido de fitoestrógenos presentes en la alfalfa (Medicago sativa) y el trébol rojo (Trifolium pratense) y evaluación de su efecto sobre el perfil hormonal en vacas Holstein en la Granja Tungüavita Paipa-Boyacá [tesis maestría]. Colombia, Tunja: Universidad Pedagógica y Tecnológica de Colombia; 2013. [ Links ]
28. Piotrowska KK, Woclawek-Potocka I, Bah MM, Piskula MK, Pilawsk W, Bober A, et al. Phytoestrogens and their metabolites inhibit the sensitivity of the bovine corpus luteum to luteotropic factors. J Reprod Dev 2006;52(1) 33-41. [ Links ]
29. Yurrita SC. Evaluation of dietary phytoestrogen exposure on growth, semen parameters, and reproductive anatomy development of growing Angus bulls [doctoral thesis]. USA, Texas: Angelo State University; 2017. [ Links ]
30. Cantero A, Sancha JL, Flores JM, Rodriguez A, Gonzalez T. Histopathological changes in the reproductive organs of Manchego ewes grazing on lucerne. J Vet Med Seri A 1996;43(6):325-330. [ Links ]
31. Pace V, Carbone K, Spirito F, Iacurto M, Terzano M, Verna M, et al. The effects of subterranean clover phytoestrogens on sheep growth, reproduction and carcass characteristics. Meat Sci 2006;74(4):616-622. [ Links ]
32. Hashem NM, El-Azrak KM, El-Din ANM, Sallam SM, Taha TA, Salem MH. Effects of Trifolium alexandrinum phytoestrogens on oestrous behaviour, ovarian activity and reproductive performance of ewes during the non-breeding season. Anim Reprod Sci 2018;196:1-8. [ Links ]
33. Pace V, Conto G, Carfi F, Chiariotti A, Catillo G. Short-and long-term effects of low estrogenic subterranean clover on ewe reproductive performance. Small Ruminant Res 2011;97(1-3):94-100. [ Links ]
34. Sierra SLA. Evaluación del efecto de los fitoestrógenos presentes en la alfalfa (Medicago sativa) sobre la calidad del semen ovino fresco y criopreservado [tesis de maestria]. Colombia, Tunja: Univesidad Pedagógica y Tecnológica de Colombia; 2015. [ Links ]
35. Ferreira-Dias G, Botelho M, Zagrajczuk A, Rebordão MR, Galvão AM, Bravo PP, et al. Coumestrol and its metabolite in mares' plasma after ingestion of phytoestrogen-rich plants: potent endocrine disruptors inducing infertility. Theriogenology 2013;80(6):684-692. [ Links ]
36. Gentao L, Yuanlin Z, Weihua C, Jie C, Zhengkang H. Effect of daidzein fed to pregnant sows on milk production and the levels of hormones in colostrum. Nanjing Agric Univ 1999;22(1):69-72. [ Links ]
37. Yuan X, Zhang B, Li L, Xiao C, Fan J, Geng M, et al. Effects of soybean isoflavones on reproductive parameters in Chinese mini-pig boars. J Anim Sci Biotechnol 2012;3(1):31. [ Links ]
38. Cardoso JR, Báo SN. Effects of chronic exposure to soy meal containing diet or soy derived isoflavones supplement on semen production and reproductive system of male rabbits. Anim Reprod Sci 2007;97(3-4):237-245. [ Links ]
39. Cardoso J, Bao S. Morphology of reproductive organs, semen quality and sexual behaviour of the male rabbit exposed to a soy-containing diet and soy-derived isoflavones during gestation and lactation. Reprod Dom Anim 2009;44(6):937-942. [ Links ]
40. Hashem NM, Abo-Elsoud MA, El-Din ANM, Kamel KI, Hassan GA. Prolonged exposure of dietary phytoestrogens on semen characteristics and reproductive performance of rabbit bucks. Dom Anim Endocrinol 2018;64:84-92. [ Links ]
41. Saleh AA, Ahmed EAM, Ebeid TA. The impact of phytoestrogen source supplementation on reproductive performance, plasma profile, yolk fatty acids and antioxidative status in aged laying hens. Reprod Dom Anim 2019;54(6):846-854. [ Links ]
42. Ni Y, Zhu Q, Zhou Z, Grossmann R, Chen J, Zhao R. Effect of dietary daidzein on egg production, shell quality, and gene expression of ER-α, GH-R, and IGF-IR in shell glands of laying hens. J Agri Food Chemis 2007;55(17):6997-7001. [ Links ]
43. Lu J, Qu L, Shen MM, Li SM, Dou TC, Hu YP, et al. Safety evaluation of daidzein in laying hens: Effects on laying performance, hatchability, egg quality, clinical blood parameters, and organ development. Poult Sci 2017;96(7):2098-2103. [ Links ]
44. Wistedt A, Ridderstråle Y, Wall H, Holm L. Effects of phytoestrogen supplementation in the feed on the shell gland of laying hens at the end of the laying period. Anim Reprod Sci 2012;133(3-4):205-213. [ Links ]
45. Arija I, Centeno C, Viveros A, Brenes A, Marzo F, Illera JC, et al. Nutritional evaluation of raw and extruded kidney bean (Phaseolus vulgaris L. var. Pinto) in chicken diets. Poult Sci 2006;85(4):635-644. [ Links ]
46. Heng D, Zhang T, Tian Y, Yu S, Liu W, Xu K, et al. Effects of dietary soybean isoflavones (SI) on reproduction in the young breeder rooster. Anim Reprod Sci 2017;177:124-131. [ Links ]
47. Cepeda VA. Disociación de respuestas estrogénicas en presencia de un extracto alcohólico de planta chilena, evaluadas en útero de rata pre-púber [tesis pregrado]. Chile, Santiago: Universidad de Chile; 2008. [ Links ]
48. Cox RI, Braden AW. The metabolism and physiological effects of phyto-oestrogens in livestock. Proc Aust Soc Anim Prod 1974;10:122. [ Links ]
49. McGarvey C, Cates PS, Brooks AN, Swanson IA, Milligan SR, Coen CW, et al. Phytoestrogens and gonadotropin-releasing hormone pulse generator activity and pituitary luteinizing hormone release in the rat. Endocrinology 2001;142(3):1202-1208. [ Links ]
50. Hughes CL, Kaldas RS, Weisinger AS, McCants CE, Basham KB. Acute and subacute effects of naturally occurring estrogens on luteinizing hormone secretion in the ovariectomized rat: Part 1. Reprod Toxicol 1991;5(2):127-132. [ Links ]
51. Goff AK. Steroid hormone modulation of prostaglandin secretion in the ruminant endometrium during the estrous cycle. Biol Reprod 2004;71(1):11-16. [ Links ]
52. McCracken JA, Custer EE, Lamsa JC. Luteolysis: a neuroendocrine-mediated event. Physiol Rev 1999;79(2):263-323. [ Links ]
53. Dubey RK, Rosselli M, Imthurn B, Keller PJ, Jackson EK. Vascular effects of environmental oestrogens: implications for reproductive and vascular health. Hum Reprod Update 2000;6(4):351-363. [ Links ]
54. Rosselli M, Reinhart K, Imthurn B, Keller PJ, Dubey RK. Cellular and biochemical mechanisms by which environmental oestrogens influence reproductive function. Hum Reprod Update 2000;6(4):332-350. [ Links ]
55. Zdunczyk S, Piskula M, Janowski T, Baranski W, Ras M. Concentrations of isoflavones in blood plasma of dairy cows with different incidence of silent heat. Bull Vet Inst Pulawy 2005;49:189-191. [ Links ]
56. Beato M. Gene regulation by steroid hormones. Cell 1989;56(3):335-344. [ Links ]
57. Romero RCM, Tarrago CMR, Muñoz MR, Arista RR, Rosado GA. Síndrome estrogénico en vacas lecheras por consumo de alfalfas con grandes cantidades de coumestrol. Vet Méx 1997;28(1):25-30. [ Links ]
58. Mustonen E, Taponen S, Andersson M, Sukura A, Katila T, Taponen J. Fertility and growth of nulliparous ewes after feeding red clover silage with high phyto-oestrogen concentrations. Animal 2014;8(10):1699-1705. [ Links ]
59. Bennetts HW, Underwood EJ, Shier FL. A specific breeding problem of sheep on subterranean clover pastures in Western Australia. Aust Vet J 1946;22(1):2-12. [ Links ]
60. Adams NR. Permanent infertility in ewes exposed to plant oestrogens. Aust Veter J 1990;67(6):197-201. [ Links ]
61. Tang BY, Adams NR. Changes in oestradiol-17β binding in the hypothalami and pituitary glands of persistently infertile ewes previously exposed to oestrogenic subterranean clover: evidence of alterations to oestradiol receptors. J Endocrinol 1978;78(2):171-177. [ Links ]
62. Arispe SA, Adams B, Adams TE. Effect of phytoestrogens on basal and GnRH-induced gonadotropin secretion. J Endocrinol 2013;219(3):243-250. [ Links ]
63. Driancourt MA, Gibson WR, Cahill LP. Follicular dynamics throughout the oestrous cycle in sheep. A review. Reprod Nutr Dev 1985;25(1A):1-15. [ Links ]
64. Cortés-Sánchez ADJ, León-Sánchez JR, Jiménez-González FJ, Díaz-Ramírez M, Villanueva-Carvajal A. Alimentos funcionales, alfalfa y fitoestrógenos. Revista Mutis 2016;6(1):28-40. [ Links ]
65. Misztal T, Wañkowska M, Górski K, Romanowicz K. Central estrogen-like effect of genistein on growth hormone secretion in the ewe. Acta Neurobiol Exp (Wars) 2007;67(4):411-419. [ Links ]
66. Lundh T. Metabolism of estrogenic isoflavones in domestic animals. Proc Soc Exp Biol Med 1995;208(1):33-39. [ Links ]
67. Takahashi O, Oishi S. Testicular toxicity of dietarily or parenterally administered bisphenol A in rats and mice. Food Chem Toxicol 2003;41(7):1035-1044. [ Links ]
68. Gutiérrez J, Castañón J. Prolapsos rectales y vaginales en cabras atribuibles al exceso de alfalfa deshidratada. En: Producción ovina y caprina. Jornadas de la Sociedad Española de Ovinotecnia y Caprinotecnia de la Universidad de Castilla-La Mancha. Castilla. 1994:239-242. [ Links ]
69. Van Weyenberg S, Sales J, Janssens G. Passage rate of digesta through the equine gastrointestinal tract: A review. Livestock Sci 2006;99(1):3-12. [ Links ]
70. Bickoff EM, Booth AN, Livingston AL, Hendrickson AP. Observations on the effect of drying on estrogenic activity of alfalfa samples of varying maturity. J Anim Sci 1960;19(3):745-753. [ Links ]
71. Gatta D, Russo C, Giuliotti L, Mannari C, Picciarelli P, Lombardi L, et al. Influence of partial replacement of soya bean meal by faba beans or peas in heavy pigs’ diet on meat quality, residual anti-nutritional factors and phytoestrogen content. Arch Anim Nutr 2013;67(3):235-247. [ Links ]
72. Groot MJ. Phyto-estrogenic activity of protein-rich feeds for pigs. RIKILT-Institute of Food Safety 2004. https://edepot.wur.nl/29220 . Accessed Mar 8, 2021. [ Links ]
73. Kuhn G, Hennig U, Kalbe C, Rehfeldt C, Ren M, Moors S, et al. Growth performance, carcass characteristics and bioavailability of isoflavones in pigs fed soy bean based diets. Arch Anim Nutr 2004;58(4):265-276. [ Links ]
74. Liu G, Chen J, Han Z. Effects of isoflavonic phytoestrogen daiazein to lactating sows. Anim Husb Vet Med 1997;29:5-7. [ Links ]
75. Ren MQ, Kuhn G, Wegner J, Nurnberg G, Chen J, Ender K. Feeding daidzein to late pregnant sows influences the estrogen receptor beta and type 1 insulin-like growth factor receptor mRNA expression in newborn piglets. J Endocrinol 2001;170(1):129-135. [ Links ]
76. Farmer C, Robertson P, Xiao C, Rehfeldt C, Kalbe C. Exogenous genistein in late gestation: effects on fetal development and sow and piglet performance. Animal. 2016;10(9):1423-1430. [ Links ]
77. Hu YJ, Gao KG, Zheng CT, Wu ZJ, Yang XF, Wang L, et al. Effect of dietary supplementation with glycitein during late pregnancy and lactation on antioxidative indices and performance of primiparous sows. J Anim Sci 2015;93(5):2246-2254. [ Links ]
78. Ratna, WN. Inhibition of estrogenic stimulation of gene expression by genistein. Life Sci 2002;71(8):865-877. [ Links ]
79. Nicholls J, Lasley BL, Nakajima ST, Setchell KD, Schneeman BO. Effects of soy consumption on gonadotropin secretion and acute pituitary responses to gonadotropin-releasing hormone in women. J Nutr 2002;132(4):708-714. [ Links ]
80. Wu N, Xu W, Cao GY, Yang YF, Yang XB, Yang XW. The blood-brain barrier permeability of lignans and malabaricones from the seeds of Myristica fragrans in the MDCK-pHaMDR cell monolayer model. Molecules 2016;21(2):134. [ Links ]
81. Kelly GE, Nelson C, Waring MA, Joannou GE, Reeder AY. Metabolites of dietary (soya) isoflavones in human urine. Clin Chim Acta 1993;223(1-2):9-22. [ Links ]
Received: November 25, 2020; Accepted: August 04, 2021
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
