











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Phycological inventories include taxonomic, ecologic and biogeographic information of species of algae, and are indispensable to recognize the biodiversity of a given region. These inventories are the result of the analysis and the integration of new and previous information, which allows comprehensive evaluations of past and current data that constitute an excellent tool to identify short and long term changes in the ecosystems (González, 1992; Huisman et al., 1998; Giangrande, 2003). Information obtained from herbaria and the revisions of bibliographic references are critical sources of data for evaluations of the biodiversity and conservation of the macroalgal flora from a certain region. These data provide an opportunity to review the state of knowledge and to examine how this knowledge of has been built over time in terms of the number of sites sampled and the number of species recognized, and to identify where there are gaps in the current inventories as far as numbers of specimens per taxon, as well as with respect to geographical and seasonal coverage (Nelson et al., 2013). Additionally, a reliable evaluation requires the taxonomic update of the species involved in order to serve as a reference for future research in the same region and to make comparisons with other regions (Serviere-Zaragoza, 1993; Candelaria-Silva, 1996; León, 1996). For this, it is always necessary to know if the sampling effort is sufficient to have a reliable inventory taking into account the rarity of the species (Ceshia et al., 2007; Capetillo et al., 2015).
The diversity has been evaluated using the taxonomic distinctness indices (Warwick & Clarke, 1995), which allows to know the variability in the composition of the different taxonomic hierarchies including the class, order, family, genus and species levels. Taxonomic distinctness is a univariate (bio) diversity index which, in its simplest form, calculates the average “distance” between all pairs of different assemblages of species; it attempts to capture phylogenetic diversity rather than simple richness of species and is more closely linked to functional diversity; it is robust to variation in sampling effort because it utilizes only simple species lists based on presence/absence data (Clarke & Warwick, 1999). In addition, according to Mistri et al. (2000), the integration and use of historic data gathered from bibliographic revisions and scientific collections that contain only presence-absence records are often used to recognize the diversity, in contrast to other indexes because the taxonomic differentiation could use independent data from different sources and it is less reliant on the sampling effort (Heino, 2005).
The taxonomic diversity indices have been used to compare different environments (e.g. marine and freshwater) (Terlizzi et al., 2005; Heino et al., 2007), regions and latitudes (e.g. polar, temperate and tropical) (Conlan et al., 2004; Miranda et al., 2005; Nicholas & Trueman, 2005; Włodarska-Kowalczuk et al., 2005; Leonard et al., 2006), being also a useful tool for assessing diversity among different sites of a certain region. Recently, these indices were used to evaluate the taxonomic diversity of different marine groups of different regions of Mexican Pacific including stony corals (Reyes-Bonilla et al., 2013), echinoderms (Rios-Jara et al., 2013), bivalves (Esqueda-González et al., 2014) and fishes (Barjau-González et al., 2012; Herrera-Valdivia et al., 2016). However, evaluations of the structure of macroalgae communities using the taxonomic distinctness to measure the variability of the biodiversity among sites by using differences between hierarchical levels are scarce (e.g. Price et al., 2006; Tonetto et al., 2016).
The tropical Mexican Pacific is one of the regions with the greatest diversity of macroalgae species in the western America (Mendoza-González & Mateo-Cid, 1998). However, previous studies are restricted to very specific taxa (Carballo et al., 2002; Senties & Dreckmann, 2014; Lopez et al., 2017), or to adjacent subtropical regions of the Gulf of California (Reyna & Rodriguez, 1996; Serviere-Zaragoza et al., 2003; Aguilar-Rosas et al., 2010; Castañeda-Fernandez de Lara et al., 2010; Mora-Valdes & Riosmena-Rodriguez, 2016; Cossete et al., 2016).
The coast of Jalisco comprise an extensive region of approximately 350 km located in the central part of the Mexican tropical Pacific, and includes a great marine biodiversity in a variety of representative tropical habitats such as rocky and sandy beaches, coral and rocky reefs, estuaries and coastal lagoons with mangrove swamps. The coast of Jalisco has been recognized for its great biodiversity of macroalgae. The first phycological records in Jalisco were those of Dawson (1949). Subsequently, the list of species increased with a series of works performed by Dawson (1951, 1953a, 1953b, 1954, 1960, 1961a, 1961b, 1962, 1963b) and later complemented by Chávez-Barrera (1980), Pedroche & González-González (1981), Enciso-Padilla et al. (1995) and Águila-Ramírez et al. (1998). In addition, the works of Serviere-Zaragoza et al. (1993, 1998), Pedroche et al. (2005, 2008) and Rodríguez et al. (2008) incorporated new records and range extensions of several species. At that moment, a total of 77 species of marine algae were reported for the coast of Jalisco; in a later work, Mendoza-González et al. (2011) updated this inventory to 165 species, this is a 214 % increase in the number of species. Since then, there are no recent inventories that integrate the macroalgal flora of the coast of Jalisco.
The present study describes an up to date taxonomic composition of macroalgae and their distribution in 19 sites of the coast of Jalisco, with an evaluation of the taxonomic categories recorded in each locality using the average Taxonomic Distinctness index (Δ+) and its variation (Λ+) in order recognize the contribution of each locality to the taxonomic diversity of this region.
Material and Methods
Study area
The state of Jalisco is located in the central-west area of the Mexican tropical Pacific (19°−21° N, 104°−106° W) and has an approximate coastal extension of 350 km (De la Lanza, 1991). The continental shelf is narrow due to its proximity to the convergent limits between the Cocos and the North America plates. This region exhibit particular oceanographic characteristics due to the circulation patterns originated by the influence of two large main current systems, the California current from the North with temperate waters, and the Equatorial countercurrent from the South with warm waters; these two great currents converge off the coasts of Nayarit and Jalisco and then flow away from the continental mass offshore to the west (Wyrtki, 1966). The physiography of most of the coastline of Jalisco is characterized by having bays and open and semi-closed coves, with sandy and rocky beaches with some small islands and islets interspaced with coastal lagoons and estuaries with mangrove swamps; in the central area there is a long sandy strip beach of approximately 50 km interrupted by coastal lagoons and several rocky beaches. Other representative tropical habitats are also present along the coastline such as coral and rocky reefs. This combination of marine environments produces a great heterogeneity that influences the structure and composition of the algae communities.
The information on macroalgae records of the coast of Jalisco was integrated from three sources:
Herbarium records
The macroalgal floral of five national herbaria was compiled: a) MEXU (Instituto de Biología, Universidad Nacional Autónoma de México); b) FCME, (Facultad de Ciencias, Universidad Nacional Autónoma de México); c) ENCB (Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional); d) GUADA (Universidad Autónoma de Guadalajara); and e) IBUG (Instituto de Botánica, Universidad de Guadalajara). A revision of material collected from the coast of Jalisco was performed in each herbarium, recording the following information: scientific name, date of collection, locality and environmental variables (intertidal level, type of substrate).
Bibliographic information
An exhaustive research of publications containing reports of species and their supra-specific categories was performed with the aim of integrating and systematizing the information on species of marine algae that have been reported in the coast of Jalisco. The research of documentary information included the revision of printed editions (books, journals, catalogs), together with the consultation of databases of both multidisciplinary and specialized electronic publications (for example ISI Web, EBSCO, ProQUEST). The use of the combination of key words (i.e. inventories, listings, distribution, macroalgae, phycoflora, algae, Jalisco) was used to refine the search depending on the language of the publication.
Collection of specimens
Sampling campaigns in 19 sites distributed along the coast of Jalisco were performed throughout all months of the year from 2000 to 2018 (19 years) (Figure 1). However, the most intensive collections were performed from March to June, during the dry season; and from September to November, during the rainy season. Collections were done along the intertidal and shallow subtidal zones (<10 m deep). Freediving (snorkel) was performed during direct collections in shallow waters (< 3 m) while SCUBA diving in deeper areas (<15 m depth). Environmental data such as wave exposure, depth, specific habitat (intertidal pool, current canal, rocky platform or cliff) and the type of substrate was associated to each specimen.

The algae collected were deposited in properly labeled plastic bags with seawater and then transported to the Laboratorio de Ecosistemas Marinos y Acuicultura (LEMA) of the Departmento de Ecología of the Universidad de Guadalajara (UdeG). One part of the samples was herborized and the other preserved in a 4 % solution of formaldehyde and seawater. Histological analysis was performed with the latter specimens for taxonomic determination, together with examination of external and internal morphology and reproductive characteristics with the aid of an optic and a stereoscope microscope. Taxonomic keys were used together with specialized bibliography such as Dawson (1944, 1947, 1953a, 1960, 1961a, 1961b, 1962), Taylor (1960), Abbott & Hollenberg (1976) and Norris and Johansen (1981), and the consultation of the database of the webpage AlgaeBase (2018). Herborized specimens were deposited in the Macroalgae collection of the Laboratorio de Ecología Molecular, Microbiología and Taxonómia (LEMITAX) of the Departamento de Ecología (UdeG).
Data Analysis
A database that compiles the composition and distribution of algae species from the coast of Jalisco was completed with an inventory of species obtained from herbaria records, documentary analysis and the collections from all sites. New macroalgae records were recognized using historic data and from new collections.
To perform the sampling effort and the Taxonomic Distinctness analysis, all the historic information and the new data from collections were grouped in 19 different sites of the north, center and south zones of Jalisco. These sites are more or less equidistant and representative of the coastal habitats of the Mexican tropical Pacific.
The sampling effort was evaluated considering the algae collected during sampling campaigns and those of historical records of the 19 selected locations of the coast of Jalisco. Species accumulation curves were done that were built with 10,000 iterations without replacement; the expected richness was calculated using the non-parametric estimators Chao 2, Jackknife 1 and Jackknife 2 by using the software EstimateS V8 (Colwell, 2006).
For the analysis of Taxonomic Distinctness (Clarke & Gorley, 2006) a presence-absence matrix for all the species recorded at 19 selected sites was completed; afterwards, a taxonomic aggregation matrix was integrated with six taxonomic levels: Division, Class, Order, Family, Genera and Species. These categories were weighted according to Warwick & Clarke (1995) in which w1 belong to species from the same genera; w2, species within the same family but different genera; w3, species within the same order but different family, and so on. The average taxonomic distinctness (Δ+) and its variation (Λ+) were estimated for each locality.
Sampling effort
All the accumulation curves of species tended towards the asymptote, indicating a good representation of the sampling effort made in the 19 selected sites of Jalisco (Figure 2). The observed species (363) represented between 65 and 74 % of the theoretical number of expected species, with an average value of 70 %. The Jackknife 2 estimator had the highest number of expected species with 403 species and, therefore, the lowest representation (66 %), while Jackknife 1 and Chao 2 had intermediate values of 359 (73 %) and 355 (74 %) ) expected species, respectively. This due to the high variability in the total number of species recorded and strong differences in species rarity among sites.
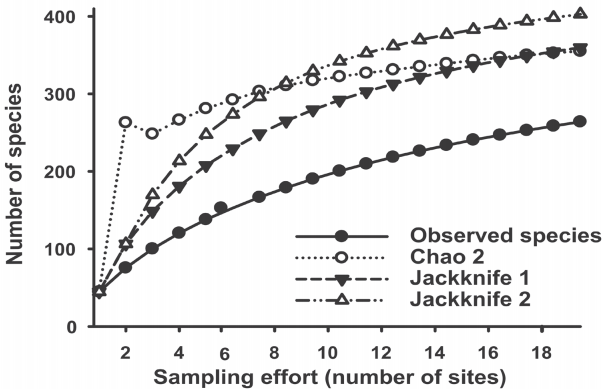
Figure 2 Accumulation curves of observed and expected macroalgae species from the coast of Jalisco, Mexico. The expected curves were estimated with the non-parametric indexes Chao 2, Jackknife 1 and Jackknife 2. The projections were constructed with 10,000 non-replacement iterations.
A high ecologic rarity (incidence) of the macroalgae was registered in Jalisco. This was evident when a more detailed revision of the observed rarity of the macroalgae species was performed, showing that an important number of species were exclusive of a single site. In fact, a total of 101 species (38 %) were unique (recorded in one site) (e.g. Asteronema breviarticulatum, Sargassum pacificum, Chondria repens, Melanothamnus bajacali, Gelidium arborescens, Amphiroa foliacea, Ulva lobata, Cladophora sericea) and, 56 (21 %) were duplicate species (recorded in two sites) (e.g. Dictyota pinnata, Ectocarpus simulans, Gracilaria tepocensis, Ulva californica, Cladophora prolifera, Bryopsis corticulans, Codium giraffa, Chlorodesmis hildebrandtii). The highest ecological rarity of the species was in the Rhodophyta (29% unique and 13 % duplicate). Also, the highest number of both unique and duplicate species was recorded in Chamela (42), Tenacatita (33), El Tamarindo (27 and Puerto Vallarta (24). In the remaining sites ecological rarity was between 1 and 21 species, with the exception of Punta Pérula with no rare species recorded.
Richness and distribution of species in the coast of Jalisco
The integration of the information gathered from three sources (revisión of herbaria material, documentary information and collections) shows a species richness of 263 macroalgae species for the coast of Jalisco, integrated in 109 genera and 61 families. The list of species is shown in Table 1, the taxonomic arrangement follows that from Silva et al. (1996) and complemented by that proposed by Wynne (1986, 2017).
Table 1 Systematic list of macroalgae species and their distribution in 19 sites of the coast of Jalisco. 1=Puerto Vallarta, 2=Yelapa, 3=Chimo, 4=Bahía Corrales, 5=Playitas, 6=Cabo Corrientes, 7=Mayto, 8=Tehuamixtle, 9= Chalacatepec, 10=Punta Pérula, 11=Chamela, 12=Isla Cocinas, 13=Careyes, 14= Tenacatita, 15=La Manzanilla, 16=El Tamarindo, 17=Cuastecomates, 18=Melaque, 19= Barra Navidad. * unique species; ** duplicates species; + new record.
| DIVISION CHLOROPHYTA |
| CLASE ULVOPHYCEAE |
| ORDEN BRYOPSIDALES |
| Familia Bryopsidaceae |
| Bryopsis corticulans Setchell 1903 (11, 14) ** |
| Bryopsis galapaguensis W.L.Taylor 1945 (14,16) ** |
| Bryopsis hypnoides J.V. Lamouroux 1809 (6, 9, 11, 18) |
| Bryopsis pennata var. minor J. Agardh 1887 (1, 9, 11, 12, 14, 16, 19) ** |
| =Bryopsis pennatula J. Agardh 1847 |
| Derbesia marina (Lyngbye) Solier 1846 (1, 11, 13) |
| Familia Caulerpaceae |
| Caulerpa chemnitzia (Esper) J.V.Lamouroux 1809 (11, 13, 14, 15, 16, 17, 19) |
| =Caulerpa peltata J.V. Lamouroux 1809 |
| Caulerpa sertularioides (S.G. Gmelin) Howe 1916 (7, 8, 12, 13, 14) |
| Familia Codiaceae |
| Codium dichotomun (Hudson) S. F. Gray 1821 (9, 11, 12, 13, 16) |
| Codium giraffa Silva 1979 (9, 11) ** |
| Codium isabelae W.R. Taylor 1945 (1, 11, 15, 16) ** |
| = Codium santamariae W.R Taylor 1945 |
| Codium picturatum Pedroche et P.C. Silva 1996 (1, 14, 15) |
| Codium setchelli N.L. Gardner 1919 (11, 12, 15) |
| Codium simulans Setchell et N.L. Gardner 1924 (1, 2, 16, 17) ** |
| =Codium cuneatum Setchell et N.L. Gardner 1924 |
| Familia Halimedaceae |
| Halimeda discoidea Decaisne 1842 (7, 8, 9, 10, 11, 12, 13, 14, 16) |
| Familia Udoteaceae |
| Chlorodesmis hildebrandtii A. Geep et E. Geep 1911 (11, 14) ** |
| ORDEN CLADOPHORALES |
| Familia Cladophoraceae |
| Chaetomorpha antennina (Bory) Kützing 1847 (1, 2, 3, 4, 6, 7, 8, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19) |
| =Chaetomorpha media (C. Agardh) Kützing 1849 |
| vChaetomorpha gracilis Kützing 1845 (19) * |
| Chaetomorpha linum (Müller) Kützing 1845 (1, 16, 18) |
| Cladophora albida (Hudson) Kützing 1843 (1) * |
| Cladophora coelothrix Kützing 1843 (11, 19) * |
| =Cladophora repens Harvey1849 |
| Cladophora laetiverens (Dillwyn) Kützing 1843 (11, 13, 16) |
| Cladophora microcladioides Collins 1909 (1, 16) ** |
| Cladophora prolifera (Roth) Kützing 1843 (1, 14) ** |
| Cladophora sericea (Hudson) Kützing 1843 (19) * |
| Cladophora vagabunda (Linneaus) van den Hoek 1963 (16, 19) ** |
| Rhizoclonium riparium (Roth) Harvey 1849 (16, 19) ** |
| =Rhizoclonium kerneri Stockmayer 1890 |
| ORDEN DASYCLADALES |
| Familia Polyphysaceae |
| Parvocaulis parvulus (Solms-Laubach) S.Berger, Fettweiss, Gleissberg, Liddle, U.Richter, Sawitzky et Zuccarello 2003 (11, 13, 14) |
| =Polyphysa parvula (Solms-Laub) Schnetter et Bula-Meyer 1982 |
| =Polyphysa parvula (Solms-Laub) Schnetter et Bula-Meyer 1982 |
| ORDEN SIPHONOCLADALES |
| Familia Siphonocladaceae |
| Boodlea composita (Harvey) Brand 1904 (15) * |
| Phyllodictyon anastomosans (Harvey) Kraft et Wynne 1996 (11) * |
| ORDEN ULVALES |
| Familia Ulvaceae |
| Ulva californica Wille 1909 (7, 11) ** |
| Ulva clathrata (Roth) C.Agardh 1811 (16, 17, 19) ** |
| =Enteromorpha crinita Nees 1820 |
| Ulva compressa Forsskål 1753 (11, 12, 14, 15, 18) |
| =Enteromorpha compressa (Linneaus) Greville 1830 |
| Ulva expansa (Setchell) Setchell et N.L. Gardner 1920 (4, 5, 7, 8, 13, 16) |
| Ulva fasciata Delile 1914 (1, 6) ** |
| Ulva flexuosa Wulfen 1803 (4, 5, 7, 8, 11, 13, 16, 17, 19) |
| =Enteromorpha flexuosa (Wulfen) J. Agardh 1883 |
| Ulva intestinalis Linnaeus 1753 (1, 8, 9, 13, 15, 16, 19) |
| =Enteromorpha intestinalis (Linneaus) Nees 1820 |
| Ulva lactuca Linnaeus 1753 (1, 2, 4, 5, 8, 9, 11, 12, 13, 14, 16, 17, 18) |
| Ulva linza Linnaeus 1753 (1, 6) ** |
| =Enteromorpha linza (Linneaus) J. Agardh 1883 |
| Ulva lobata (Kützing) Setchell et N.L. Gardner 1920 (13) * |
| Ulva prolifera O.F.Müller 1778 (1, 6, 11, 16) |
| =Enteromorpha prolifera (Müller) J. Agardh 1883 |
| =Enteromorpha salina Kützing 1845 |
| Ulva rigida C. Agardh 1823 (11, 13, 14, 15, 17, 18, 19) |
| Ulva taeniata (Setchell) Setchell et N.L. Gardner 1920 (6, 8, 11, 13, 14, 16, 18) |
| =Ulva dactylifera Setchell et N.L. Gardner 1920 |
| Familia Ulvellaceae |
| Phaeophila dendroides (P. Crouan et H. Crouan) Batters 1902 (11) * |
| Ulvella viridis (Reinke) R.Nielsen, C.J.O’Kelly et B.Wysor in R.Nielsen et al. 2013 (19)* |
| =Acrochaete viridis (Reinke) R.Nielsen 1979 |
| DIVISION OCHROPHYTA |
| CLASE PHAEOPHYCEAE |
| ORDEN DICTYOTALES |
| Familia Dictyotaceae |
| Dictyopteris delicatula J.V. Lamouroux 1809 (14, 15, 16) |
| Dictyota bartayresiana J.V. Lamouroux 1809 (10, 13, 14) |
| Dictyota ciliolata Kützing 1859 (10, 11, 13) |
| Dictyota crenulata J. Agardh 1847 (1, 2, 3, 11, 13, 14, 15, 18) |
| Dictyota dichotoma (Hudson) J.V. Lamouroux 1809 (8, 10, 12, 16) |
| Dictyota dichotoma var. intricata (C. Agardh) Greville 1830 (13, 14, 17) |
| Dictyota implexa (Desfontaines) J.V.Lamouroux 1809 (1, 10, 13, 14, 16) |
| =Dictyota divaricata J.V. Lamouroux 1809 |
| Dictyota flabellata (Collins) Setchell et N.L. Gardner 1924 (17, 18) ** |
| Dictyota friabilis Setchell 1926 (4, 11, 17) |
| =Dictyota pfaffii Schnetter 1972 |
| Dictyota pinnata (E.Y.Dawson) I.Hörnig, R.Schnetter et Prud’homme van Reine 1993 (11, 13, 17) |
| =Dilophus pinnatus E.Y. Dawson 1950 |
| Lobophora variegata (J.V. Lamouroux) Womersley ex Oliveira 1977 (13, 15) |
| ** |
| Padina caulescens (Thivy) W.R. Taylor 1945 (1, 9, 10, 11, 12, 13, 14, 15, 16, 19) |
| Padina concrescens Thivy 1945 (8, 11) ** |
| Padina crispata (Thivy) W.R. Taylor 1945 (1, 2, 4, 9, 11, 13, 15, 17, 18) |
| Padina durvillaei Bory 1827 (1, 2, 3, 4, 6, 11, 13, 14, 16) |
| Padina gymnospora (Kützing) Sonder 1871 (1, 2, 4, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19) |
| =Padina vickersiae Hoyt 1920 |
| Padina mexicana E.Y. Dawson 1944 (1, 5, 7, 12, 14) |
| Padina pavonica J.V. Lamouroux 1816 (8, 17,18) |
| Zonaria farlowii Setchell et N.L. Gardner 1924 (2, 10, 11, 12, 13, 17, 18) |
| ORDEN ECTOCARPALES |
| Familia Chordariaceae |
| Compsonema serpens Setchell et N.L. Gardner 1922 (1) * |
| Hecatonema streblonematoides (Setchell et N.L.Gardner) Loiseaux 1970 (1, 11, 15) |
| v=Compsonema secundum Setchell et N.L. Gardner 1922 |
| Hecatonema terminale (Kützing) Kylin 1937 (14) * |
| Streblonema penetrale Setchell et N.L. Gardner 1922 (18) * |
| Familia Acinetosporaceae |
| Feldmannia mitchelliae (Harvey) H.-S.Kim 2010 (13, 15, 17, 19) |
| =Hincksia mitchelliae (Harvey) P.C.Silva 1987 |
| =Giffordia mitchelliae (Harvey) Hamel 1939 |
| Familia Ectocarpaceae |
| Ectocarpus parvus (Saunders) Hollenberg 1971 (1, 11, 18) |
| Ectocarpus rallsiae Vickers 1905 (13) * |
| =Hincksia rallsiae (Vickers) P.C.Silva 1987 |
| Ectocarpus siliculosus (Dillwyn) Lyngbye 1819 (11, 14) ** |
| Ectocarpus simulans Setchell et N.L. Gardner 1922 (1, 11) ** |
| Hincksia saundersii (Setchell et N.L. Gardner) P.C.Silva in P.C. Silva, E.G. Meñez et Moe 1987 (16) * |
| =Giffordia saundersii (Setchell et N.L. Gardner) Hollenberg et Abbott 1966 |
| ORDEN FUCALES |
| Familia Sargassaceae |
| Sargassum agardhianum Farlow 1889 (1) * |
| Sargassum alternato-pinnatum Yamada 1942 (1, 2) ** |
| =Sargasum asymmetricum E.Y. Dawson 1944 |
| Sargassum brandegeei Setchell et N.L. Gardner 1924 (11) * |
| Sargassum horridum Setchell et N.L. Gardner 1924 (11) * |
| Sargassum howellii Setchell 1937 (1, 2, 3, 4, 7, 8, 10, 11, 12, 14, 15, 16, 17, 18, 19) |
| Sargassum liebmanii J. Agardh 1847 (1, 2, 3, 4, 6, 8, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19) |
| Sargassum pacificum Bory 1828 (1) * |
| ORDEN RALFSIALES |
| Familiy Hapalospongidiaceae |
| Hapalospongidion gelatinosum Saunders 1899 (18) * |
| Familia Neoralfsiaceae |
| Neoralfsia hancockii (E.Y.Dawson) D.Leon-Alvarez et M.L.Nunez-Reséndiz in Leon-Alvarez et al. 2014 (13, 14, 15) |
| = Ralfsia hancockii E.Y.Dawson 1944 Familia Ralfsiaceae |
| Ralfsia confusa Hollenberg 1969 (1, 4, 11, 14) |
| Ralfsia fungiformis (Gunner) Setchell et N.L. Gardner 1924 (11, 18) ** |
| Ralfsia hesperia Setchell et N.L. Gardner 1924 (1, 4, 11, 18) |
| Ralfsia pacifica Hollenberg 1944 (1, 17, 18) |
| ORDEN SCYTOSIPHONALES |
| Familia Chnoosporaceae |
| Chnoospora minima (Hering) Papenfuss 1956 (1, 2, 3, 4, 9, 10,11, 12, 13, 14, 15, 16, 17, 18) |
| =Chnoospora pacifica J. Agardh 1847 |
| Familia Scytosiphonaceae |
| Colpomenia sinuosa (Roth) Derbés et Solier 1856 (1, 16) ** |
| Petalonia fascia (O.F.Müller) Kuntze 1898 (14) * |
| =Ralfsia californica Setchell et N.L. Gardner 1924 |
| ORDEN SCYTOTHAMNALES |
| Familia Bachelotiaceae |
| Bachelotia antillarum (Grunow) Gerloff 1959 (14) * |
| Familia Asteronemataceae |
| Asteronema breviarticulatum (J. Agardh) Oriques et Bouzon 2000 (11, 15, 17, 18, 19)* |
| = Ectocarpus breviarticulatus J. Agardh 1847 (11)* |
| ORDEN SPHACELARIALES |
| Familia Lithodermateceae |
| Pseudolithoderma nigrum Hollenberg 1969 (17) * |
| Familia Sphacelariaceae |
| Sphacelaria californica Sauvageau 1925 (1) * |
| Sphacelaria tribuloides Meneghini 1840 (16) * |
| =Sphacelaria mexicana W.R.Taylor 1945 |
| Sphacelaria novae-hollandiae Sonder 1845 (11, 13) ** |
| Sphacelaria rigidula Kützing 1843 (1, 2, 11, 19) |
| DIVISON RHODOPHYTA |
| CLASE BANGIOPHYCEAE |
| ORDEN BANGIALES |
| Familia Bangiaceae |
| Bangia fuscopurpurea (Dillwyn) Lyngbye 1819 (11, 14) ** |
| Piropya raulaguilarii (5)* + |
| CLASE FLORIDEOPHYCEAE |
| ORDEN ACROCHAETIALES |
| Familia Acrochaetiaceae |
| Acrochaetium hancockii (E.Y.Dawson) Papenfuss 1945 (11, 15) ** |
| Acrochaetium microscopicum (Nägeli ex Kützing) Nägeli 1858 (14, 15) ** |
| ORDEN AHNFELTIALES |
| Familia Ahnfeltiaceae |
| Ahnfeltia plicata (Hudson) Fries 1835 (6, 12, 13, 18) |
| Ahnfeltia svensonii W.R. Taylor 1945 (1, 2, 6, 11, 13, 14, 15, 17, 18) |
| ORDEN BONNEMAISONIALES |
| Familia Bonnemaisoniaceae |
| Asparagopsis taxiformis (Delile) Trevis 1845 (11, 14) ** |
| ORDEN CERAMIALES |
| Familia Callithamniaceae |
| Aglaothamnion endovagum (Setchell et N.L.Gardner) I.A.Abbott 1972 (11) * |
| =Callithamnion endovagum Setchell et N.L.Gardner 1924 |
| Callithamnion rupiculum C.L.Anderson 1894 (11) * |
| Familia Ceramiaceae |
| Antithamnionella breviramosa (E.Y. Dawson) Womersley et Bailey 1970 (1) * |
| Antithamnionella elegans (Berthold) J.H.Price et D.M.John 1986 (14, 19) ** |
| Centroceras clavulatum (C. Agardh) Montagne 1846 (1, 2, 4, 5, 6, 7, 8, 10, 11, 12, 13, 14, 15, 16, 17, 18) |
| Ceramium affine Setchell et N.L. Gardner 1930 (14) * |
| Ceramium camouii E.Y Dawson 1944 (19) * |
| Ceramium caudatum Setchell et N.L. Gardner 1924 (14, 16) ** |
| Ceramium hamatispinum E.Y. Dawson 1950 (13, 15, 19) |
| Ceramium zacae Setchell et N.L. Gardner 1937 (15, 16, 19) |
| Gayliella flaccida (Harvey ex Kützing) T.O.Cho et L.J.McIvor 2008 (1, 4, 11, 13, 14, 15, 16, 17, 18, 19) |
| =Ceramium flaccidum (Kützing) Ardissone 1871 |
| Gayliella fimbriata (Setchell & N.L. Gardner) T.O.Cho et S.M.Boo 2008 (11, 14, 15, 18) |
| =Ceramium fimbriatum Setchell et N.L. Gardner 1924 |
| Gayliella taylorii (E.Y. Dawson) T.O.Cho et S.M.Boo 2008 (5, 8, 11, 12, 13, 14, 15) |
| =Ceramium taylorii E.Y. Dawson 1950 |
| Familia Dasyaceae |
| Dasya sinicola var. abyssicola (E.Y. Dawson) E.Y. Dawson 1963 (11) * |
| Familia Delesseriaceae |
| Taenioma perpusillum (J. Agardh) J. Agardh 1848 (11, 15) ** |
| Familia Rhodomelaceae |
| Bostrychia moritziana (Sonder ex Kützing) J.Agardh 1863 (19) * |
| Chondria acrorhizophora Setchell et N.L.Gardner 1924 (2, 6, 15, 19) |
| =Chondria californica (Collins) Kylin 1941 |
| Chondria arcuata Hollenberg 1945 (1, 2, 16) |
| Chondria repens Borgesen1924 (15) * |
| Herposiphonia littoralis Hollenberg 1970 (11, 16) ** |
| Herposiphonia secunda (C.Agardh) Ambronn 1880 (16, 17) ** |
| Herposiphonia subdisticha Okamura1899 (14) * |
| Herposiphonia tenella (C. Agardh) Ambronn 1880 (1, 2, 11, 16) |
| Herposiphonia verticillata (Harvey) Kylin 1925 (1) * |
| Laurencia clarionensis Setchell et N.L. Gardner 1937 (15) * |
| Laurencia decidua E.Y. Dawson 1954 (11, 13) ** |
| Laurencia gardnerii Hollenberg 1943 (17) * |
| Laurencia hancockii E.Y.Dawson 1944 (14, 16) ** |
| Laurencia humilis Setchell et N.L Gardner 1930 (14, 16) ** |
| Laurencia lajolla E.Y. Dawson 1958 (11) * |
| Melanothamnus bajacali (Hollenberg) Díaz-Tapia et Maggs in Díaz-Tapia et al. 2017 (16) * |
| =Polysiphonia bajacali Hollenberg 1961 |
| Melanothamnus johnstonii (Setchell et N.L.Gardner) Díaz-Tapia et Maggs in Díaz-Tapia et al. 2017 (14, 16) ** |
| =Polysiphonia johnstonii Setchell et N.L. Gardner 1924 |
| Melanothamnus savatieri (Hariot) Díaz-Tapia et Maggs in Díaz-Tapia et al. 2017 (15) * |
| =Polysiphonia japonica var. savatieri (Hariot) H.Y.Yoon 1986 |
| Melanothamnus simplex (Hollenberg) Díaz-Tapia et Maggs 2017 (1, 14, 15, 17, 18, 19) |
| =Neosiphonia simplex (Hollenberg) Y.P.Lee 2008 |
| =Polysiphonia simplex Hollenberg 1942 |
| Melanothamnus sphaerocarpus (Borgesen) Díaz-Tapia et Maggs in Díaz-Tapia et al. 2017 (13) * |
| =Neosiphonia sphaerocarpa (Borgesen) M.S.Kim et I.K.Lee 1999 |
| Murrayellopsis dawsonii Post 1962 (14, 16) ** |
| Osmundea sinicola (Setchell et N.L. Gardner) K.W.Nam 1944 (13) * |
| Polysiphonia confusa Hollenberg 1961 (14, 16) ** |
| Polysiphonia hendryi N.L. Gardner 1927 (16) * |
| Polysiphonia mollis Hooker et Harvey 1947 (19) * |
| Polysiphonia pacifica Hollenberg 1942 (16, 17, 18) |
| Polysiphonia sertularioides (Grateloup) J. Agardh 1863 (14) * |
| Polysiphonia subtilissima Montagne 1840 (19) * |
| Symphyocladia dendroidea (Montagne) Savoie et G.W.Saunders 2016 (14, 16) ** |
| =Pterosiphonia dendroidea (Montagne) Falkenberg 1901 |
| Tayloriella dictyurus (J. Agardh) Kylin 1941 (1, 2, 3, 4, 6, 11, 13, 14, 15, 16, 17) |
| Familia Wrangeliaceae |
| Anotrichium tenue (C.Agardh) Nägeli 1862 (11) * |
| Pleonosporium globuliferum Levring 1941 (11) * |
| ORDEN COLACONEMATALES |
| Familia Colaconemataceae |
| Colaconema daviesii (Dillwyn) Stegenga 1985 (1, 2, 15, 19) |
| =Acrochaetium daviesii (Dillwyn) Nägeli 1862 |
| Colaconema hypneae (Borgesen) A.A.Santos et C.W.N.Moura 2010 (14, 15, 19) |
| =Acrochaetium hypneae (Borgesen) Borgesen 1915 |
| Colaconema savianum (Meneghini) R.Nielsen 1994 (11, 13, 19) |
| =Acrochaetium savianum (Meneghini) Nägeli 1862 |
| Grania pectinata (Kylin) Athanasiadis 2016 (14) * |
| =Colaconema pectinatum (Kylin) J.T.Harper et G.W.Saunders 2002 |
| ORDEN CORALLINALES |
| Familia Corallinaceae |
| Amphiroa beauvoisii J.V.Lamouroux 1816 (1, 2, 3, 11, 12, 13, 14, 15, 16, 17, 18) |
| =Amphiroa drouetii E.Y.Dawson 1953 |
| =Amphiroa mexicana W.R. Taylor 1945 |
| Amphiroa dimorpha Lemoine 1929 (1, 2, 3, 5, 7, 8, 9, 11, 12, 13, 14, 16, 17, 18) |
| Amphiroa foliacea J.V. Lamouroux 1824 (3) * |
| Amphiroa franciscana W.R. Taylor 1945 (16) * |
| Amphiroa misakiensis Yendo 1902 (1, 11, 13, 14, 16, 17, 18) |
| = Amphiroa brevianceps E.Y.Dawson 1953 Amphiroa polymorpha Lemoine 1929 (1) * |
| Amphiroa rigida J.V. Lamouroux 1816 (1, 2, 16) |
| Amphiroa subcylindrica E.Y. Dawson 1946 (18) * |
| Amphiroa taylorii E.Y. Dawson 1934 (2, 16) ** |
| Amphiroa valonioides Yendo 1902 (1, 13, 18) |
| Bossiella californica (Decaisne) Silva 1957 (17) * |
| Crusticorallina muricata (Foslie) P.W.Gabrielson, Martone, K.R.Hind et C.P.Jensen in Hind et al. 2016 (2) * |
| = Lithophyllum lichenare Mason 1953 |
| Fosliella paschalis (Lemoine) Setchell et N.L. Gardner 1930 (2) * |
| Hydrolithon farinosum (J.V. Lamouroux) Penrose et Chamberlain 1993 (1, 2, 11, 14) |
| =Fosliella farinosa (J.V. Lamouroux) Howe 1920 |
| Jania adhaerens J.V. Lamouroux 1816 (1, 2, 4, 10, 11, 12, 13, 14, 16, 17, 19) |
| Jania capillacea Harvey 1853 (18) * |
| Jania longiarthra E.Y. Dawson 1949 (11, 16) ** |
| Jania pacifica Areschoug 1852 (1, 2, 3, 4, 11, 13, 14, 15, 16, 18, 19) |
| =Jania mexicana W.R. Taylor 1945 |
| = Titanoderma corallinae (P.Crouan et H.Crouan) Woelkerling, Y.M. Chamberlain et P.C.Silva 1985 |
| Lithophyllum decussatum (J.Ellis et Solander) Philippi 1837 (17) * |
| Lithophyllum dispar (Foslie) Foslie 1909 (1, 15, 17) |
| =Tenarea dispar (Foslie) Adey 1970 Lithophyllum imitans Foslie 1909 (1, 2, 18) |
| Lithophyllum proboscideum (Foslie) Foslie 1900 (18) * |
| Lithophyllum stictaeforme (Areschoug) Hauck 1877 (11) * |
| Melobesia polystromatica E.Y. Dawson 1954 (14) * |
| Melobesia membranacea (Esper) J.V. Lamouroux1812 (11, 14) ** |
| Neogoniolithon setchellii (Foslie) Adey 1970 (1) * |
| Neogoniolithon trichotomum (Heydrich) Setchell et N.L. Gardner 1943 (1, 14) ** |
| Familia Hydrolithaceae |
| Pneophyllum fragile Kützing 1843 (13) * |
| Pneophyllum nicholsii (Setchell et L.R. Mason) P.C.Silva et P.W.Gabrielson 2004 (1, 2, 6) |
| = Heteroderma nicholsii Setchell et N.L. Gardner 1943 |
| Familia Lithophyllaceae |
| Titanoderma pustulatum (J.V.Lamouroux) Nägeli in Nägeli et Cramer 1858 (14, 15) ** |
| =Titanoderma pustulatum var. confine (P.Crouan et H.Crouan) Y.M.Chamberlain 1991 |
| Familia Lithothamniaceae |
| Phymatolithon lenormandii (Areschoug) Adey 1966 (15) * |
| Familia Porolithaceae |
| Dawsoniolithon conicum (E.Y.Dawson) Caragnano, Foetisch, Maneveldt et Payri in Caragnano et al 2018 (14, 15) ** |
| =Pneophyllum conicum (E.Y. Dawson) Keats,Y.M.Chamberlain et M.Baba 1997 |
| Harveylithon rupestre (Foslie) A.Rösler, Perfectti, V.Peña et J.C.Braga 2016 (11) * |
| =Hydrolithon rupestre (Foslie) Penrose 1996 |
| Harveylithon samoënse (Foslie) A.Rösler, Perfectti, V.Peña et J.C.Braga 2016 (13) * |
| =Hydrolithon samoënse (Foslie) Keats et Y.M.Chamberlain 1994 |
| Porolithon onkodes (Heydrich) Foslie 1909 (14) * |
| =Hydrolithon onkodes (Heydrich) Penrose et Woelkerling 1992 |
| Familia Spongitaceae |
| Spongites decipiens (Foslie) Y.M. Chamberlain 1993 (1, 14, 18, 15) |
| =Hydrolithon decipiens (Foslie) Adey 1970 |
| =Lithophyllum decipiens (Foslie) Foslie 1900 |
| ORDEN HALYMENIALES |
| Familia Halymeniaceae |
| Grateloupia clarionensis (Setchell et N.L. Gardner) S.Kawaguchi & H.W.Wang 2001 (14, 19) ** |
| Grateloupia doryphora (Montagne) Howe 1914 (4, 6, 11, 13, 14, 16) |
| = Grateloupia multiphylla E.Y. Dawson 1954 |
| Grateloupia filicina (J.V. Lamouroux) C. Agardh 1822 (1, 16) ** |
| Grateloupia hancockii E.Y Dawson 1944 (11) * |
| Grateloupia huertana Mateo-Cid, Mendoza-González et Gavio 2005 (11, 14, 15) |
| Grateloupia howeii Setchell et N.L. Gardner 1924 (1, 2, 4, 11) |
| Grateloupia prolongata J. Agardh 1847 (13, 16) ** |
| Grateloupia versicolor (J. Agardh) J. Agardh 1851 (1, 11, 13, 15, 19) |
| Grateloupia sp. (5, 7, 8) |
| Prionitis mexicana E.Y Dawson 1944 (11) * |
| Familia Endocladiaceae |
| Gloiopeltis furcata (Postels et Ruprecht) J. Agardh 1851 (17) * |
| Familia Peyssonneliaceae |
| Cruoriella fissurata E.Y.Dawson 1953 (14) * |
| Peyssonnelia orientalis (Weber-van Bosse) Cormaci et G.Furnari 1987 (1, 6, 15) |
| Peyssonnelia pacifica Kylin 1925 (15) * |
| ORDEN ERYTHROPELTIDALES |
| Familia Erythrotrichiaceae |
| Erythrotrichia carnea (Dillwyn) J. Agardh 1883 (3, 13, 14, 15, 18) |
| Erythrotrichia tetraseriata N.L. Gardner 1927 (11, 13) ** |
| Sahlingia subintegra (Rosenvinge) Kornmann 1989 (1, 2, 14) |
| Smithora naiadum (Anderson) Hollenberg 1959 (6) * |
| ORDEN GELIDIALES |
| Familia Gelidiaceae |
| Gelidium arborescens N.L. Gardner 1927 (16) * |
| Gelidium crinale (Turner) Gaillon1825 (13) * |
| Gelidium mcnabbianum (E.Y Dawson) B.Santelices 1998 (19) * |
| Gelidium microdentatum E.Y Dawson 1960 (4, 6, 9, 11) |
| Gelidium pusillum (Stackhouse) Le Jolis 1863 (1, 2, 6, 8, 9, 10, 11, 12, 13, 14, 17, 18) |
| Gelidium sclerophyllum W.R. Taylor 1945 (1, 3, 6, 9, 13, 14, 15, 17, 18, 19) |
| Gelidium sp. (5, 7, 8) |
| Familia Gelidiellaceae |
| Gelidiella acerosa (Forkssal) J. Feldmann et Hamel 1934 (1) * |
| Gelidiella hancokii E.Y.Dawson1944 (19) * |
| Parviphycus antipae (Celan) B. Santelices 2004 (15) * |
| Familia Pterocladiaceae |
| Pterocladiella caloglossoides (M.Howe) Santelices 1998 (1, 18) ** |
| =Pterocladia caloglossoides (M.Howe) E.Y. Dawson 1953 |
| Pterocladiella capillacea (S.G.Gmelin) Santelices et Hommersan 1997 (4) * |
| =Pterocladia capillacea (S.G.Gmelin) Bornet 1876 |
| Pterocladiella media (E.Y. Dawson) Boo & Miller in Boo et al. 2006 (2) * |
| =Pterocladia media E.Y. Dawson 1958 |
| ORDEN GIGARTINALES |
| Familia Gigartinaceae |
| Chondracanthus harveyanus (Kützing) Guiry in Hommersand, Guiry, Fredericq et Leister 1993 (6) * |
| =Gigartina harveyana (Kützing) Setchell et Gardner 1933 |
| Chondracanthus spinosus (Kützing) Guiry in Hommersand, Guiry, Fredericq et Leister 1993 (6, 17) ** |
| =Gigartina asperifolia J. Agardh 1899 |
| =Gigartina spinosa (Kützing) Harvey 1853 |
| Mazzaella leptorhynchos (J.Agardh) Leister in Hommersand et al. 1993 (16, 17) ** |
| =Gigartina leptorhynchos J. Agardh 1885 |
| Mazzaella volans (C.Agardh) Fredericq in Hommersand et al 1993 (17) * |
| =Gigartina volans (C. Agardh) J. Agardh 1846 Familia Hypneaceae |
| Hypnea johnstonii Setchell et Gardner 1924 (1, 3, 4, 6, 11, 13, 15, 17) |
| Hypnea pannosa J. Agardh 1847 (1, 9, 11, 12, 14, 16, 17, 18) |
| Hypnea spinella (C. Agardh) Kützing 1847 (1, 5, 8, 11, 13, 14, 15, 16, 17, 18, 19) |
| Hypnea valentiae (Turner) Montagne 1841 (17, 18) ** |
| Familia Solieriaceae |
| Solieria filiformis (Kützing) P.W.Gabrielson 1985 (5, 7) ** + |
| Wurdemannia miniata (Duby) Feldmann et Hamel 1934 (14) * |
| Familia Phyllophoraceae |
| Ahnfeltiopsis concinna (J.Agardh) Silva et De Cew 1992 (5, 11, 12, 13, 14, 16, 18) |
| = Ahnfeltia concinna J. Agardh 1847 |
| Ahnfeltiopsis serenei (E.Y. Dawson) Masuda 1993 (11) * |
| Besa leptophylla (J.Agardh) M.S.Calderon et K.A.Miller in Calderon et al 2016 (1, 2, 4, 6, 16) |
| =Gymnogongrus leptophyllus J. Agardh 1876 |
| Gymnogongrus crustiformis E.Y. Dawson 1961 (16) * |
| Gymnogongrus johnstonii (Setchell et N.L. Gardner) E.Y. Dawson 1954 (4, 9, 13, 14, 17, 18) |
| Gymnogongrus martinensis Setchell et N.L.Gardner 1937 (11) * |
| Gymnogongrus platyphyllus N.L. Gardner 1927 (16) * |
| ORDEN GRACILARIALES |
| Familia Gracilariaceae |
| Gracilaria crispata Setchell et N.L. Gardner 1924 (2, 6, 17, 18) |
| Gracilaria pachydermatica Setchell et N.L.Gardner 1924 (1) * |
| Gracilaria ramisecunda E.Y.Dawson 1949 (11) * |
| Gracilaria rubrimembra E.Y.Dawson 1949 (11) * |
| Gracilaria spinigera E.Y. Dawson 1949 (11, 13, 16) |
| Gracilaria symmetrica E.Y. Dawson 1949 (11) * |
| Gracilaria tepocensis (E.Y. Dawson) E.Y. Dawson 1944 (6, 13) ** |
| Gracilaria textorii (Suringar) De Toni 1876 (13, 16, 17) |
| Gracilariopsis costaricensis E.Y. Dawson 1964 (14) * |
| ORDEN HILDENBRANDIALES |
| Familia Hildenbrandiaceae |
| Hildenbrandia dawsonii (Ardré) Hollenberg 1971 (6) * |
| Hildenbrandia rubra (Sommerfelt) Meneghini 1841 (1, 11) ** |
| =Hildenbrandia prototypus Nardo 1834 |
| ORDEN NEMALIALES |
| Familia Galaxauraceae |
| Tricleocarpa cylindrica (Ellis et Solander) Huisman et Borowitzka 1990 (13) * |
| Familia Liagoraceae |
| Dermonema virens (J.Agardh) Pedroche et Ávila Ortíz 1996 (11, 13, 14, 15, 17, 18) |
| = Dermonema frappierii (Montagne et Millardet) Borgesen 1942 Izziella orientalis (J.Agardh) Huisman et Schils 2002 (15) * |
| Familia Scinaiaceae |
| Scinaia confusa (Setchell) Huisman 1985 (18) * |
| =Gloiophloea confusa (Setchell) Levring 1914 |
| CLASE STYLONEMATOPHYCEAE |
| ORDEN STYLONEMATALES |
| Familia Stylonemataceae |
| Stylonema alsidii (Zanardini) Drew 1956 (1, 11, 19) |
| ORDEN RHODYMENIALES |
| Familia Champiaceae |
| Champia parvula (C. Agardh) Harvey 1853 (14, 15, 17, 18) |
| Familia Lomentariaceae |
| Ceratodictyon tenue (Setchell et N.L.Gardner) J.N.Norris 2014 (11) * |
| = Gelidiopsis tenuis Setchell et N.L.Gardner 1924 |
| Ceratodictyon variabile (J.Agardh) R.E.Norris 1987 (1, 11, 13, 14, 15, 18) |
| >=Gelidiopsis variabilis (J. Agardh) Schmitz 1895 |
| Familia Rhodymeniaceae |
| Botryocladia uvarioides E.Y. Dawson 1944 (1) * |
| Halichrysis irregularis (Kützing) A. Millar 2005 (11) * |
| Rhodymenia californica Kylin 1931 (16) * |
Three divisions of macroalgae were recorded, Rhodophyta (red algae), Ochrophyta (brown algae) and Chlorophyta (green algae); the red algae had the highest number of all taxonomic levels from Class to species (Table 2).
Table 2 Number of species, genera, families, orders and classes of the three divisions of macroalgae recorded in the Jalisco coast.
| Division | Species | Genera | Families | Orders | Classes |
|---|---|---|---|---|---|
| Rhodophyta | 167 | 72 | 37 | 17 | 4 |
| Ochrophyta | 52 | 22 | 14 | 7 | 1 |
| Chlorophyta | 44 | 17 | 10 | 5 | 1 |
The distribution of the species was very variable along the coast of Jalisco. The largest number of records was found in Puerto Vallarta (84) in the North, Chamela (104) and, Careyes (71) in the center, and, Tenacatita (86) and, El Tamarindo (74) in the South (Table 3). However, the areas with lower richness (14-27 species), are also from Chimo to Tehuamixtle in the North, and also from Chalacatepec to Isla Cocinas (except for Chamela) in the center. The southern sites had intermediate (54-62) to high (71-86) species richness.
Two species, Jania tenella and Chaetomorpha antennina had the widest distribution along the coast of Jalisco; these species were recorded in 17 sites, followed by Sargasum liebmanii and, Centroceras clavulatum (16 sites). Fifteen species were present along the coastline (10 to 15 sites): Ulva lactuca, Halimeda discoidea, Chnoospora minima, Padina caulescens, Padina gymnospora, Sargassum howellii, Hypnea spinella, Tayloriella dictyurus, Gelidium pusillum, Gayliella flaccida, Gelidium sclerophyllum, Amphiroa beauvoisii, Amphiroa dimorpha, Jania adhaerens and, Jania pacifica. However, the macroalgae of Jalisco are characterized by their restricted distribution due to ecological rarity, these include 101 species (38 %) of 38 different families recorded in only one site (unique species), and, 56 species (21 %) from 28 families recorded in two sites (duplicate species). The species Ectocarpus siliculosus, Cladophoropsis sundanensis and, Phyllodictyon anastomosans, among others, have been recognized as annual algae according to their life cycle, this means that they frequently live only during the most favorable season of the year (Mate-Cid & Mendoza-González, 2012). This may one of the reasons why these species has a low incidence (ecological rarity) in the coast of Jalisco.
Taxonomic Distinctness
The values of the average taxonomic distinctness (∆+) of all sites fell within the probability channel (e.g. within the confidence intervals of 95 %, p>0.05), with the exception of Bahía Chamela (Figure 3). These sites had a greater contribution to the mean taxonomic diversity of macroalgae of the coast of Jalisco. The sites that contributed the most were Cuastecomates, La Manzanilla, El Tamarindo and, Tenacatita. On the other hand, values of the variation of taxonomic distinctness (Λ+) showed that most sites are significantly representative of the macroalgae assemblage, only four sites, Punta Pérula, Chimo, Yelapa and, El Tamarindo, fell out of the 95 % probability channel. This indicates that most of the sites along the coast of Jalisco, have a good representation of the different taxonomic categories of macroalgae (Figure 3; Table 3).
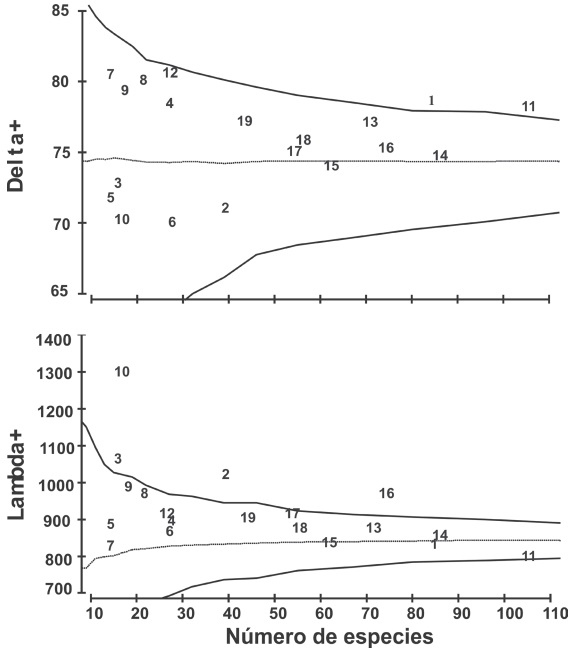
Figure 3 Average taxonomic distinctness ( +) and the variation of the taxonomic distinctness (Λ +) of the macroalgae from the coast of Jalisco, Mexico. The solid line shows the 95% confidence intervals and the dashed line the values of + & Λ +. The statistical significance of + and Λ + was assessed using 1,000 permutations. See Table 3 for symbology.
Table 3 Number of macroalgae taxa recorded in 19 sites of the coast of Jalisco, Mexico. sites are arranged from north to south.
| Site / Taxa | Species | Genera | Families | Order | Classes | Division |
|---|---|---|---|---|---|---|
| 1. Puerto Vallarta | 84 | 45 | 34 | 21 | 4 | 3 |
| 2. Yelapa | 39 | 29 | 20 | 16 | 3 | 3 |
| 3. Chimo | 15 | 11 | 9 | 9 | 3 | 3 |
| 4. Bahía Corrales | 26 | 16 | 14 | 11 | 3 | 3 |
| 5. Playitas | 14 | 11 | 10 | 8 | 4 | 3 |
| 6. Cabo Corrientes | 27 | 19 | 17 | 13 | 3 | 3 |
| 7. Mayto | 14 | 12 | 11 | 10 | 3 | 3 |
| 8. Tehuamixtle | 21 | 13 | 11 | 10 | 3 | 3 |
| 9. Chalacatepec | 18 | 11 | 10 | 7 | 3 | 3 |
| 10. Punta Pérula | 16 | 10 | 8 | 8 | 3 | 3 |
| 11. Chamela | 104 | 53 | 40 | 27 | 5 | 3 |
| 12. Isla Cocinas | 26 | 18 | 15 | 11 | 3 | 3 |
| 13. Careyes | 71 | 39 | 30 | 21 | 4 | 3 |
| 14. Tenacatita | 86 | 57 | 39 | 23 | 5 | 3 |
| 15. La Manzanilla | 62 | 41 | 32 | 21 | 4 | 3 |
| 16. El Tamarindo | 74 | 36 | 22 | 15 | 4 | 3 |
| 17. Cuastecomates | 54 | 32 | 23 | 19 | 4 | 3 |
| 18. Melaque | 55 | 32 | 26 | 19 | 4 | 3 |
| 19. Barra Navidad | 44 | 26 | 20 | 16 | 4 | 3 |
New records of species
The exploration of new sites as well as the increase of sampling effort at sites with single or very few references, allowed to identify two new records for the coast of Jalisco, Solieria filiformis (Kützing) of the Family Solieriaceae, collected from October to May in Playitas and Mayto; and, Piropya raulaguilarii, a unique species of the Family Bangiaceae, collected only in April in Playitas. Both macroalgae were found in the low intertidal of rocky beaches.
Discussion
The completion of an updated inventory of the phycoflora from the coast of Jalisco, integrated from different sources of information, allowed to increase the number of species to 263; this is more than 100 % compared to the last inventory made by Pedroche et al. (2005, 2008), who reported 152 species for the coast of Jalisco. It is important to point out that the most recent inventory was made by Mendoza-González et al. (2011), who recorded the same number of species (152) only in the southern coast of Jalisco. Although many records were found in several specific reports and herbarium, any of them represent true compilations or complete inventories for the entire coast of Jalisco. However, all these were taken into account as part of the documented phycoflora of this region and included in our inventory. The only two new records reported here were found during our own samplings performed in the coast of Jalisco. It is also important to note that with this inventory the species richness from the coast of Jalisco is greater than that found in other adjacent areas, such as the coast of Nayarit with 184 species (Mateo-Cid & Mendoza-González, 1992; Serviere et al., 1993), located in the northern limit of the Mexican tropical Pacific; and the coast of Colima with 125 (Mateo-Cid & Mendoza- González 1991), located towards the south of Jalisco; this indicates the great contribution of the coast of Jalisco to the biodiversity of macroalgae. The species of Jalisco represents 60 % of the total number of species (444) compiled by Pedroche & Sentíes (2003) for the Tropical Mexican Pacific. Finally, comparisons with other recent inventories from the Gulf of California (63 in Norris, 2010; and 124 in Mora-Valdez & Riosmena-Rodriguez, 2016), suggest that the inventory of 263 species here presented has a good representation of the diversity of algae of the Mexican Pacific.
The Rhodophyta dominated the taxonomic diversity of all macroalgae recorded in Jalisco, with the highest number of species (167) and all taxa levels from Class to Genus. These algae grow successfully as epizoic, epiphytic or epilithic on the surface of animals, other algae or rocks with a higher coverage and diversity in tropical marine environments (Mendoza-González et al., 2011). Other studies performed in the Tropical Mexican Pacific have recorded a lower diversity of red algae: Mateo-Cid & Mendoza-González (1992) reported 69 species for the southern coast of Nayarit; Mateo-Cid & Mendoza- González (2002), 93 species for the coast of Guerrero; Mateo-Cid & Mendoza- González (2003), 87 species from Oaxaca; and Dreckmann et al. (2006), only 29 species from Chiapas.
The analysis of the distribution of the macroalgae along the northern, center and southern coast of Jalisco showed some areas with different taxonomic diversity. The sites with higher number of species are protected rocky beaches within bays which are semi-protected from waves and currents, with small barriers or patches of coral reefs. The structural complexity of the rocky and coralline substrate is a key factor for de recruitment and growth of the algal populations. The crevices, hollows and other irregularities of the hard substrate create microenvironments for the settlement of propagules and provide protection to depredation and physical perturbation (Lüning, 1990; Dudley & D’Antonio, 1991). On the other hand, an extensive area of low species richness of macroalgae located in the northern part from Chimo to Tehuamixtle, is characterized by long sandy beaches intercalated by few rocky beaches. Similarly, some sites of the central zone with few species have few suitable places (for example, rocky substrates) for the development of the phycoflora.
Many species of the subtidal environment are found in mixed substrates of rocks, skeletons of dead coral and sand, mainly in shallow areas (<20 m). Although the distribution of the macroalgae showed a greater abundance and number of species in locations with rocky reefs and scattered coral colonies, there were also macroalgae associated with other substrates, such as boulders, pebbles, gravel and sand. For example, some species of the families Cladophoraceae (Cladophora spp), Caulerpaceae (Caulerpa chemnitzia, C. sertularioides) and Halimedaceae (Halimeda discoidea) were common on sandy substrates and gravel. The rocky and coral reefs have the highest biodiversity of invertebrates (scleractina, gorgonians, polychaete worms, echinoderms, molluscs) and fish, with a significant presence of associated macroalgae.
In the coast of Jalisco, there are two protected marine areas: “Chamela-Cuixmala” and “Islas de la Bahía de Chamela”, both in the central region, where four sites of the present study are included (Punta Pérula, Chamela, Isla Cocinas and Careyes). Both areas have the main objective of preserving their biodiversity and ecological balance (CONANP, 2008), and they offer benefits to the inhabitants of the region, generating activities such as fishing and recreation, including diving, which are relevant activities due to their natural scenic beauty.
The historical records of the different groups of organisms are difficult to interpret when they are analyzed in terms of abundance, since they are affected by factors such as the sampling method and the sampling effort; sample size, records of environmental and ecological variables (Clarke & Warwick, 1998, Leonard et al., 2006, Ceshia et al., 2007). The biodiversity measured with the taxonomic distinctness (Δ + and Λ +), has been successfully used in other marine taxonomic groups with information based on the presence/absence of Echinodermata species (: (Ríos-Jara, et al. 2013);); bivalve molluscs (Esqueda-González, et al., 2014); macroalgae, macroinvertebrates and fish (García et al., 2014). The records on the specific richness of macroalgae obtained from the different sources used in our study allowed the identification of four sites that contributed the most to the taxonomic diversity of the marine algae of Jalisco (Figure 3); they were within the probability channel of 95 % of the average taxonomic distinction (Δ +). These four sites (Cuastecomates, La Manzanilla, El Tamarindo and Tenacatita) are located in the southern coastal area and have been historically documented as those with the greatest contribution to the diversity of macroalgae in the region.
The high number of unique species recorded contributed to a great extent, to the phycofloristic mosaic observed in the present study, representing 38% of the total. A characteristic of this group of unique species is that most of them are filamentous organisms, less visible and with a reduced habit size, such as the organisms of Orders Ceramiales, Rhodymeniales, Ectocarpales and Cladophorales (Table 1). These attributes, along with the characteristics of each species, could have been the reason why they were recorded only once. A significant number of these species have been reported as epiphytes of other algae such as Erythrotrichia carnea, Bangia fuscopurpurea, Acrochaetium hancockii, Colaconema daviesii, Asteronema breviarticulatum; therefore, their observation and record could have been occasional and conditioned to the collection and careful observation of the host algae, which in most cases were foliar algae such as Sargassum, or flabelated such as Padina.
It is important to note that the sites in which the greatest numbers of unique species were recorded in the southern coast; historically, this is the region with the largest number of phycological studies. This could be an important factor to increase the records of unique species; since this was not observed in the sites of the northern coast which have been recently explored (Tehuamixtle, Mayto, Playitas). However, it is precisely in this region where two new records for the phycoflora of Jalisco were found: Solieria filiformis and Piropya raulaguilarii, which shows that this must be a high priority region for future studies on marine flora and fauna.
The present study demonstrates that the phycoflora of the coast of Jalisco is diverse and it also contains characteristic elements of the Mexican tropical Pacific. However, more studies are needed for a better understanding of the structure of the macroalgae assemblages of Jalisco, with estimations of coverage and, finally, the biomass of species with a potential exploitation. The study of the different sites along the coast of Jalisco was of great relevance to integrate a more complete taxonomic inventory of the macroalgae, which is essential to identify strategies for the conservation and use of these ecosystems. At this time, this has great relevance because there are several proposals of tourism development for the coast of Jalisco. Finally, the exploitation of macroalgae with different purposes (for example, food, pharmacology) is under development in Mexico, with the species of shallow intertidal and subtidal environments being the main potential resources due to their accessibility and abundance (Carrillo-Domínguez et al., 2002; Hernández-Carmona et al., 2012). In the Mexican tropical Pacific, some common species, for example, Sargassum liebmannii (Ochrophyta), Caulerpa sertularioides (Chlorophyta) and Spyridia filamentosa (Rhodophyta), have shown good biological activity and potential use in food and pharmacology (Radulovich et al., 2015; Garcia-Granados et al., 2016; Landa-Cansigno et al., 2017). However, any proposal for its use must consider evaluations of exploitable biomass to guarantee the sustainable use of these resources.
Conclusions
Based on the updated inventory integrated from new and previous information, a total of 263 species of macroalgae from 19 sites of the coast of Jalisco, Mexico, including new records of species, were recognized. Macroalgae of Jalisco are characterized by a high ecological rarity (incidence), with 38 % of unique species (recorded only in one site) and 21 % of duplicated species (in two site). This inventory highlights the contribution of this region to the diversity of macroalgae of the Mexican tropical Pacific with at least 60 % of the species commonly found in subtidal, intertidal and shallow environments. The Jalisco coast is one of the regions with the highest species richness of the Eastern tropical Pacific (Pedroche et al., 2005, 2008), including Central America (Pedroche & Sentíes, 2003a, Pedroche & Sentíes, 2003b, Pedroche et al., 2003).

