











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkHighlights:
Passive restoration led to reactivation of biogeochemical cycle in the forest.
Decomposition rates indicated high returns of organic matter and nutrients to the soil.
Phosphorus had low returns via leaf litter and low release from standing litter.
Cordia alba had the best performance from a biogeochemical cycling perspective.
Introduction
The tropical dry forest represents 42 % of tropical ecosystems worldwide; however, it is poorly understood especially in terms of dynamics and functioning (Becknell, Kuceck, & Powers, 2012; Rai, Singh, Ghosal, & Singh, 2016). At present, increasing human pressure, together with livestock farming and climate change, generate deterioration of this ecosystem (Becknell et al., 2012; Carbonó & García, 2010; Ferreira & Vieira, 2017).
Colombia has about 1 200 of the original 80 000 km2 of tropical dry forest (Flórez-Flórez, León-Peláez, Osorio-Vega, & Restrepo-Llano, 2013), distributed mainly on the Caribbean coast, where the situation is worrisome in cities and populated centers, as representative samples of the ecosystem as part of the urban green infrastructure are practically nonexistent.
In cities, forests influence the improvement of the perception of well-being of their inhabitants (Madureira, Andresen, & Monteiro, 2011), fulfilling important ecological, environmental, social and economic functions (Ferrini & Fini, 2011; Soares et al., 2011). Forest degradation implies the loss of environmental services, so ecological restoration activities should aim not only at the recovery of structure and composition, but also at the recovery of functionality (León & Osorio, 2014).
Understanding the dynamics of urban forests is important for determining their resilience, a key aspect in the design of urban green infrastructure to mitigate problems such as climate change and environmental degradation in cities (Colding, 2011).
Ecological processes such as nutrient cycling, via leaf litter production and decomposition, determine key trajectories in the resilience process and support of ecosystem services as a result of ecological restoration activities (Castellanos, Blanco, León, & Chavez, 2019; León & Osorio, 2014). These processes lead to the renewal of nutrients in the soil and the subsequent reestablishment of biogeochemical cycles. (Castellanos-Barliza & León, 2011), aspects that have special significance in the functioning of green areas and forests in urban environments.
This study shows the results of the monitoring of nutrient cycling in an urban tropical dry forest in a successional stage, located on land that was used for agricultural production (10 years of abandonment). The main objective was to characterize the accumulation and decomposition of organic materials and the flow of nutrients from standing litter. Based on the study of these processes, the potential use of three species (Albizia niopoides Spruce ex Benth., Cordia alba [Jacq.] Roem. & Schult. and Machaerium milleflorum Dugand G. A.) was determined for future active restoration activities.
Materials and Methods
Study area
The study was carried out in an 11-year-old secondary dry forest fragment, located in the city of Santa Marta, Colombia (11° 13' N y 74° 11' W; 21 m). Mean annual temperature is 27 °C with mean annual precipitation of 608.8 mm (Montes-Correa et al., 2015). The dominant tree species are A. niopoides, C. alba and M. milleflorum with stand-level densities of 50, 85 and 36 individuals∙ha-1, respectively. The soils are alkaline (pH 8.2) with high contents of organic matter (4.0 %) and exchangeable bases. The forest fragment developed spontaneously after the abandonment of agricultural activities, mainly cotton crops, representing a passive restoration strategy. A more detailed description of the characteristics of the study site can be found in Castellanos-Barliza, León-Peláez, Armenta-Martínez, Barranco-Pérez, and Caicedo-Ruíz (2018).
Monitoring and classification of standing litter
A total of fourteen samples were taken from the leaf litter layer accumulated on the soil surface or standing litter (O horizon), each with an area of 0.5 m2. The plant material samples were taken to the Biogeochemistry laboratory of the Universidad Nacional de Colombia, Medellín, where they were dried in an oven at 65 °C, until constant dry weight. The following fractions were separated for each sample: leaves of A. niopoides (LA), leaves of C. alba (LC), leaves of M. milleflorum (LM), leaves of other species (LOS), reproductive material (RM), woody material (WM) and other remains (OR). Leaf material of the dominant species (LA, LC and LM) was ground for ash determination in a 600 °C muffle. Subsequently, elemental concentrations of nitrogen (N) and phosphorus (P) were obtained by micro-Kjeldahl and acid digestion (L-ascorbic), respectively. Calcium (Ca), magnesium (Mg) and potassium (K) were determined by dry digestion and atomic absorption reading (Perkin Elmer Instruments 2380, USA).
Leaf litter decomposition and nutrient fluxes
The rate of leaf litter decomposition in standing litter was determined by calculating the decomposition coefficient k j = A/(F + A) (Jenny, Gessel, & Bingham, 1949); where, A is the amount of leaf litter falling annually from the canopy to the ground and F is the amount of leaf litter accumulated on the ground.
The values of leaf litter production (A) used for the calculations were obtained from leaf litter traps installed in the forest fragment, the results have been published in Castellanos-Barliza et al. (2018). The mean residence time (MRT, years) of leaf litter and nutrients were calculated as the inverse of the coefficient k j (MRT = 1/k j ) (León, González, & Gallardo, 2011). The potential nutrient return rate (PRR), via leaf litter, was calculated by multiplying the leaf dry mass by the concentration of each nutrient. The actual return rate of each nutrient (ARR) was calculated by multiplying the PRR by the respective decomposition constant k j . The amount of each nutrient stored in standing litter leaves, nutrient retention by standing litter (NRSL), was calculated by multiplying the leaf dry mass of the leaves by the concentration of the nutrient in them. Finally, for each nutrient, an effective rate of return (ERR) was obtained by the coefficient between PRR and NRSL. ERR values close to 1 indicate that the amounts of nutrients reaching the soil surface annually, via leaf litter, are similar to those that accumulate in litter layer. Low ERR values indicate nutrient accumulation in standing litter (León et al., 2011).
Data analysis
Data was subjected to analysis of variance (ANOVA) and Tukey's comparison of means (P = 0.05), to determine differences between the dry mass values of the standing litter fractions, as well as their potential return via fine leaf litter and nutrient concentrations. Previously, heteroscedasticity and homogeneity of variances (normality test) were tested. The analyses were performed using STATGRAPHICS Centurion version 16.2 (Statpoint Technologies, 2013).
Results
Leaf litter accumulation and decomposition
Table 1 shows the results of the analysis of leaf litter decomposition and accumulation on the soil of an urban forest fragment. Total standing litter accounted for 8.3 Mg∙ha-1, in which woody materials (39 %) and other residues (34 %) accounted for the greatest participation, while reproductive material had a smaller participation (6 %). Leaf litter from standing litter represented 20 % of the total; M. milleflorum and A. niopoides had similar participation (12 and 13 %, respectively), while C. alba only represented 3 %. Leaves of other species were predominant with a value of 1.2 Mg∙ha∙year-1, representing 71 % of the leaves in the standing litter (Figure 1).
Table 1 Standing leaf litter and other fractions in an urban dry forest fragment in Santa Marta, Colombia.
| Variable | LC | LM | LA | LOS | TSLL | WM | RM | OR | TSL |
|---|---|---|---|---|---|---|---|---|---|
| Standing litter (kg∙ha-1) | 51 ± 3.8 a | 206 ± 15.9 a | 224 ± 12.9 a | 1 190 ± 43.5 bd | 1 671 ± 42.3 b | 3 265 ± 69.5 c | 535 ± 19.7 d | 2 878 ± 104.80 c | 8 349 ± 101.37 |
| CV (%) | 148 | 155 | 116 | 73 | 51 | 43 | 74 | 73 | 24 |
| Fraction (%) | 3.07 | 12.33 | 13.39 | 71.21 | 20.01 | 39.11 | 6.41 | 34.47 | 100 |
CV: coefficient of variation. Different letters indicate statistically significant difference according to Tukey's tes (P < 0.05). ± Standard deviation of the mean (n = 14). LC = Cordia alba, LM = Machaerium milleflorum, LA = Albizia niopoides, LOS = leaves of other species, TSLL = total standing leaf litter, WM: woody material, RM: reproductive material, OR: other residues, TSL: total standing litter.
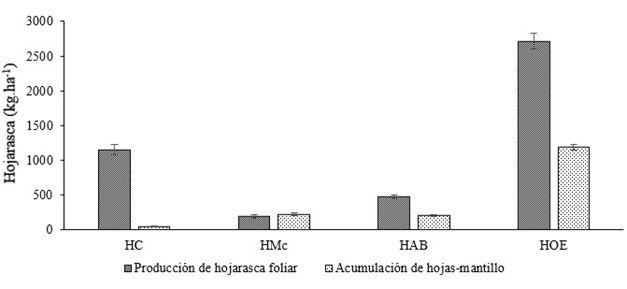
Figure 1 Annual leaf litter production and accumulation of leaves in standing litter for the studied species (LC: Cordia alba, LM: Machaerium milleflorum, LA: Albizia niopoides, LOS: leaves of other species) in an urban tropical dry forest fragment in Santa Marta, Colombia.
Leaf litter production was higher than leaf accumulation in standing litter, except for M. milleflorum leaves (Figure 1). Based on Table 2, the greatest difference between production and accumulation was determined for C. alba, which was reflected in the constant k j implying higher decomposition rate (k j = 0.96) and lower mean residence time (MRT = 1.04 years). Of the three species studied, A. niopoides showed the lowest decomposition rate (k j = 0.46) and the longest residence time (2.15 years). For the total standing litter, the constant k j = 0.50 implies a time of 2.01 years for total decomposition to be verified.
Table 2 Decomposition and mean residence time (MRT) of standing litter fractions of the species studied in an urban dry forest fragment in Santa Marta, Colombia.
| Fractions | A(kg∙ha-1∙year-1) | F(kg∙ha-1∙year-1) | kj | MRT (years) |
|---|---|---|---|---|
| LC | 1 155.11 | 51.23 | 0.96 | 1.04 |
| LM | 475.09 | 206.02 | 0.7 | 1.43 |
| LA | 193.42 | 223.77 | 0.46 | 2.16 |
| LOS | 2 716.76 | 1 189.63 | 0.7 | 1.44 |
| TSLL | 4 540.37 | 1 670.65 | 0.73 | 1.37 |
| TSL | 8 248.00 | 8 349.19 | 0.5 | 2.01 |
A: anual leaf litter production, F: amount of standing litter, k j : Jenny's decomposition constant (Jenny et al., 1949), LC: leaves of Cordia alba, LM: leaves of Machaerium milleflorum, LA: leaves of Albizia niopoides, LOS: leaves of other species, TSLL: total standing leaf litter, TSL: total standing litter.
Nutrient release from standing leaf litter
According to Table 3, N was the element with higher values in standing litter (NRSL) for all species (LM = 4.16 kg∙ha-1∙year-1; LA = 4.95 kg∙ha-1∙year-1; LC = 0.60 kg∙ha-1∙year-1), showing, in turn, the lowest values of the release constant k j . In contrast, P had the lowest accumulation in standing leaf litter (LM = 0.08 kg∙ha-1∙year-1; LA = 0.07 kg∙ha-1∙year-1; LC = 0.07 kg∙ha-1∙year-1). For cations, accumulation in standing litter followed the decreasing sequence Ca > Mg > K in all species. According to the ERR values, species had higher efficiency in returning cations than N and P. The highest efficiency was for C. alba followed by M. milleflorum and A. niopoides.
Table 3 Indices of nutrient return, retention and release processes via standing leaf litter for each of the species studied in an urban dry forest fragment in Santa Marta, Colombia.
| Indices | N | P | Ca | Mg | K |
|---|---|---|---|---|---|
| (kg∙ha-1∙year-1) | |||||
| Machaerium milleflorum | |||||
| PRR | 7.74 | 0.31 | 18.67 | 1.78 | 2.64 |
| NRSL | 4.16 | 0.08 | 1.83 | 0.1 | 0.16 |
| kj | 0.65 | 0.79 | 0.91 | 0.95 | 0.94 |
| MRT | 1.54 | 1.26 | 1.1 | 1.06 | 1.06 |
| ARR | 5.04 | 0.25 | 17 | 1.68 | 2.48 |
| ERR | 1.86 | 3.8 | 10.18 | 17.25 | 16 |
| Albizia niopoides | |||||
| PRR | 4.81 | 0.16 | 3.64 | 0.37 | 1.01 |
| NRSL | 4.95 | 0.07 | 1.55 | 0.11 | 0.13 |
| kj | 0.49 | 0.71 | 0.7 | 0.77 | 0.88 |
| MRT | 2.03 | 1.42 | 1.42 | 1.31 | 1.13 |
| ARR | 2.37 | 0.11 | 2.55 | 0.28 | 0.89 |
| ERR | 0.97 | 2.4 | 2.35 | 3.27 | 7.54 |
| Cordia alba | |||||
| PRR | 14.66 | 1.29 | 36.54 | 17.33 | 16.28 |
| NRSL | 0.6 | 0.07 | 0.5 | 0.05 | 0.09 |
| kj | 0.96 | 0.95 | 0.99 | 1 | 0.99 |
| MRT | 1.04 | 1.06 | 1.01 | 1 | 1.01 |
| ARR | 14.09 | 1.22 | 36.04 | 17.29 | 16.19 |
| ERR | 24.37 | 18.01 | 72.37 | 377.63 | 177.33 |
PRR: potential nutrient return rate via leaf litter, NRSL: nutrient retention in standing leaf litter, ARR:actual nutrient return rate, k j : Jenny's decomposition constant, MRT: mean residence time (years), ERR: effective rate of return.
Discussion
The values of litter accumulation and decomposition, after the abandonment of productive activities, show a visible reactivation of the biogeochemical cycle in this tropical dry forest ecosystem in successional stage.
Total standing litter was high (8.3 Mg∙ha-1∙year-1) when compared to values reported for dry forests in early successional stage. Restrepo, Flórez, León, and Osorio (2013) evaluated successional patches of “mosquero” (Croton leptostachyus Kunth) and neem (Azadirachta indica A. Juss) plantations of ages younger than the fragment studied here (six years). The study aimed to determine how the reactivation of the biogeochemical cycle led, via fine leaf litter contributions, to soil improvement; litter accumulation values were much lower (C. leptostachyus: 369 kg∙ha-1∙year-1 and A. indica: 451 kg∙ha-1∙year-1) to those of the present study. These values of standing litter (expressed as dry mass) are the result of the relationship between production and decomposition rate of organic residues contributed by the forest canopy (León et al., 2011), so it is better to use decomposition rates to interpret the dynamics of biogeochemical cycling more accurately at the ecosystem level. Thus, in the study by Restrepo et al. (2013), k j values for total standing litter indicated higher decomposition rates in C. leptostachyus successional patches than in A. indica plantations (k j = 0.71 vs. 0.55). These values of total standing litter decomposition are higher than those obtained in this study (k j = 0.5), which has a mean residence time of about two years (Table 2) that is lower than that reported for tropical dry forests in Mexico (MRT = 2.79 years Campo & Vázquez-Yanes, 2006). Despite the low k j value in the present study, organic matter returns to the soil from total standing litter decomposition, in absolute terms, are higher than those reported by Restrepo et al. (2013); fine leaf litter production values are very high (8.3 Mg∙ha-1∙year-1), compared to those of A. indica (0.6 Mg∙ha-1∙año-1) and C. leptostachyus (0.9 Mg∙ha-1∙year-1).
As an integral part of standing litter, the study of leaf fraction is of special interest in the biogeochemical cycle because it represents the major component of fine litter and is relatively easy to decompose, thus representing a fundamental transfer pathway for organic matter and nutrients (Dutta & Agrawal, 2001; Norgrove & Hauser, 2000). The results of both standing litter dynamics and, particularly, leaf fraction are relevant for this dry forest fragment, as they indicate a high return of organic matter (k j = 0.73; MRT = 1.37 years, Table 2). In this ecosystem, leaves collectively represent 20 % of the standing litter, whereas, according to Castellanos-Barliza et al. (2018), the leaves represented 55 % (4.7 Mg∙ha-1) of leaf litter shed from the canopy. This suggests an intense fragmentation of leaves in standing littter towards the fraction other residues, whose participation is very high (35 %, Table 1). Regarding the values of the decomposition constant k j of this fraction (k j = 0.73) and the annual leaf litterfall just indicated, organic matter returns from decomposition of leaf materials would represent about 3.4 Mg∙ha-1∙year-1 in this ecosystem. This value is higher than those projected as decomposition returns from k j values for A. indica plantations (33 kg C∙ha-1∙year-1) and for C. leptostachyus patches (72 kg C∙ha-1∙year-1) in Colombia. Based on k j values of litter-bags and leaf litterfall values, organic matter returns projected for dryland silvopastoral systems in Colombia by Martínez, Cajas, León, and Osorio (2014) were much lower (448 to 770 kg∙ha-1∙year-1).
From the analysis at the species level (Table 2), the rate of leaf decomposition followed the decreasing sequence C. alba > M. milleflorum > A. niopoides. Since this order is also found in litter production, the species C. alba, with visibly higher values, has a high potential for inclusion in restoration activities and, in general, for reforestation of land in dry climates. León and Osorio (2014) suggest that species to be included in ecological restoration activities and, specially, in the rehabilitation of degraded lands should exhibit good adaptability and high growth rates, as well as high production of rapidly decomposing leaf litter.
Nutrient release from standing litter
N was the nutrient with the highest accumulation in litter layer, as a result of the low values of the release constant k j (Table 3); however, high rates of N return through canopy leaf litter (PRR) produced in turn high absolute values of actual return (ARR), which were only lower than those of Ca, the element for which the absolute maximums were determined in the three species. Only for C. alba, the ARR values of Mg and K were higher than those of N. The opposite situation occurred with P, a nutrient that was released in minimum quantity from the leaves in standing litter, reaching absolute minimums in ARR among nutrients studied. Low levels of P released from standing litter (ARR) are based on a low return through leaf litter for the species studied (PRR), result of a low foliar concentration of the nutrient (0.07 a 0.11 %, Castellanos-Barliza et al., 2018).
A low level of P in leaf litter provides a severe restriction on plant growth and, specifically, on the activity of soil microorganisms, because of the high requirements of the nutrient (Swift, Heal, & Anderson, 1979). Thus, a critical value of 12 has been proposed for the N/P ratio of leaf litter (Aerts, 1997); in tropical environments, this value represents some degree of P scarcity for decomposer organisms, since in fungal and bacterial cells such a ratio is around 10 to 15. N/P values for the three species (A. niopoides = 24, M. milleflorum = 23, C. alba = 11; Castellanos-Barliza et al., 2018) suggest that only C. alba produces leaf litter favorable for soil microbial activity and, with it, for nutrient recycling in the ecosystem. In fact, the lower value of the N/P ratio was reflected in a faster decomposition of C. alba leaves (Table 2) with respect to the other two species with higher N/P values.
In conclusion, in this tropical dry forest fragment, the abandonment of agricultural activities (10 years) and the consequent advance of natural succession allowed the reactivation of the biogeochemical cycle, based on the production and decomposition of fine leaf litter. The relatively high rates of litter production and decomposition in this ecosystem were probably influenced by agricultural field management practices prior to abandonment. However, a severe ecosystem-level constraint on P availability was evident, with low amounts of return through leaf litter and release from standing litter (PRR, ARR, k j ), as well as low IER values. Of the three species, only C. alba had favorable performance, despite this restriction to ecosystem functioning. This species, in addition to producing a high amount of easily decomposable leaf litter, puts into circulation larger amounts of P that do not bioaccumulate, but are released for the most part, as demonstrated by the maximum values of IER.
Conclusions
Restoration processes in tropical dry forests will surely be favored by an adequate supply of nutrients, mainly P, which, although present in the soil, is precipitated and therefore poorly bioavailable for both plants and microorganisms. In fact, it has been stated that vegetation included in restoration models usually must overcome the low availability of P in the soil to achieve satisfactory growth and development. The results suggest that Cordia alba is suitable for incorporation in restoration activities. Furthermore, it is important to carry out more studies of this nature that allow the identification of a greater number of species to increase the success of restoration initiatives in these environments.

