











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
The genus Agave s. l. currently includes ca. 265 species (Thiede 2020). Approximately, 226 of these species correspond to Agave s. s., of which 153 (68 %) are native to Mexico, the center of origin of the group, 40 (18 %) are native to the Caribbean, 21 (9 %) to the United States, 13 (6 %) to Central America, six (3 %) to northern South America, and nine (4 %) are only known from cultivation (García-Mendoza 2002, Vázquez-García et al. 2007a, García-Mendoza et al. 2019, Thiede 2020). On a broader scale, Western Mexico or Nueva Galicia (Rzedowski & McVaugh 1966) is considered to have the highest concentration of Agave species in the world (Vázquez-García et al. 2007b, c, 2016, Cuevas-Guzmán et al. 2010). On a smaller scale, the Valle de Tehuacán and its surroundings are the center of highest species diversity (Reichenbacher 1985, García-Mendoza & Galván-Villanueva 1995, Thiede 2020, Eguiarte et al. 2021).
Ever since the publication of the monumental Agave monograph nearly four decades ago (Gentry 1982), the number of Agave s. s. species has increased considerably (53 spp.), six of the 30 sections in the subgenera Agave and Littaea (Thiede et al. 2019) include 70 % of the newly described species: 10 in A. sect. Heteracanthae Salm-Dyck (Galván-Villanueva & Hernández-Sandoval 2002, García-Mendoza et al. 2007, Vázquez-García et al. 2007a, González-Elizondo et al. 2011), Hernández-Sandoval & Magallán-Hernández 2015, García-Mendoza et al. 2019, Hernández-Vera et al. 2019, Starr & Davis 2019, Starr 2020), followed by eight in A. sect. Juncineae Salm-Dyck (Zamudio-Ruiz & Sánchez-Martínez 1995, García-Mendoza & Martínez-Salas 1998, Carrillo-Reyes et al. 2003, Cabral-Cordero et al. 2007, Galván-Villanueva & Zamudio-Ruiz 2013, García-Mendoza & Chávez-Rendón 2013, Starr et al. 2018, García-Morales et al. 2019), seven in A. sect. Ditepalae Hochstätter (Hodgson & Slauson 1995, Hodgson 2001, Vázquez-García et al. 2012, Hodgson & Salywon 2013, Hodgson et al. 2018, García-Mendoza et al. 2019), five in A. sect. Marmoratae (A.Berger) Thiede & Gideon F.Sm. (Cházaro-Basáñez et al. 2005, Vázquez-García et al. 2013), five in A. sect. Micracanthae Salm-Dyck (Cházaro-Basáñez 1995, Cházaro-Basáñez et al. 2010, Cházaro-Basáñez & Vázquez-García 2013, Arzaba-Villalba et al. 2018, Beutelspacher & Hernández-Najarro 2021), and four in A. sect. Inermes Salm-Dyck, including the one here proposed as new (Cházaro-Basáñez et al. 2006, Vázquez-García et al. 2007a, b, Cuevas-Guzmán et al. 2012). Most of the newly described species (73 %) are from western Mexico (13 spp.), Oaxaca-Puebla-Guerrero (12), eastern Mexico (8), and northern Mexico (6).
The populations from western Oaxaca here described as a new species, A. rosalesii, were first placed in A. ellemeetiana K.Koch (1865), a species with stemless rosettes of few soft and flat, entire leaves without a terminal spine and yellow flowers with a very short corolla tube. Agave ellemeetiana was published from plants cultivated in Europe (Thiede 2014); its habitats were unknown for more than 150 years until its confirmed occurrence in central Veracruz and northern Oaxaca (Cházaro-Basáñez et al. 2012, Thiede 2014).
Further populations ascribed to A. ellemeetiana, but differing in its finely dentate leaf margins, were reported from the Putla / Tlaxiaco area in western Oaxaca (Köhres 2008). Thiede (2014) applied the forgotten name A. ellemeetiana var. subdentataTrelease (1914) to these plants; this name and the plants were unknown to Gentry (1982). Thiede (2014) had only photographs and small cultivated plants raised from field-collected seeds at hand, from which he preserved several leaves as neotype. He wanted to leave Trelease's name as it was published, pending field studies assessing the characteristics, variability and differences of the plants from the Putla / Tlaxiaco area to decide whether to recognize them as subspecies ofA. ellemeetianaor a species in its own right. However, on behalf of one of the reviewers, Thiede (2014) had to publish the new combination A. ellemeetiana K.Koch ssp. subdentata (Trel.) Thiede, thus raising Treleases's name to subspecific rank.
A molecular genetic study using AFLP markers (Gil-Vega et al. 2007) displays a clear separation between A. ellemeetiana (ssp. ellemeetiana) and the Putla populations corresponding to A. ellemeetiana ssp. subdentata (Thiede 2014), grouping the latter with A. pedunculifera (from Guerrero), A. vilmoriniana and A. bracteosa. Similarly, A. attenuata (ssp. attenuata) groups with A. garciae-mendozae Galván & L.Hern., clearly separate from A. pedunculifera (from Tlacotepec, Guerrero). In the light of this genetic evidence, A. pedunculifera can be maintained as a species distinct from A. attenuata and not as a subspecies of the latter, as proposed by Ullrich (2006).
Recent field studies unveiled that the flowers of the populations from the Putla District, Oaxaca, exhibit a distinct corolla tube and thus do not correspond to A. sect. Choritepalae (type species: A. ellemeetiana) with a very short corolla tube, but instead to A. sect. Inermes (lectotype species: A. attenuata) with a distinct corolla tube. Thus, there is no reason to keep the Putla populations as a subspecies of A. ellemeetiana, instead, we here propose a distinct new species placed in the Inermes section. A closer look at the flowers shows additional differences in flower shape, color and length, as well as in the color of the filaments and styles. Furthermore, we highlight the habitat and ecological differences among similar taxa in both the Choritepalae and Inermes sections.
Agave section Inermes currently includes 11 species, of which two are known from cultivation only: A. gilbertii A. Berger and A. spicata Cav. The rest is confined to the Pacific slopes of Mexico from Sonora to Oaxaca (Figure 1): Agave attenuata Salm-Dyck, A. chazaroi A.Vázquez & O.M. Valencia, A. chrysoglossa I.M. Johnst., A. manantlanicola Cuevas & Santana-Mich., A. nizandensis Cutak, A. ocahui Gentry, A. pedunculifera Trel., A. vazquezgarciae Cházaro & J.A.Lomelí, and A. vilmoriniana A. Berger (Gentry 1982, McVaugh 1989, Cházaro-Basáñez et al. 2006, Vázquez-García et al. 2007a,b, Cuevas-Guzmán et al. 2012, Thiede 2020). Molecular studies revealed that A. sect. Inermes is not monophyletic (Gil-Vega et al. 2007, Hernández-Vera et al. 2007, Jiménez-Barron et al. 2020), but its long corolla tube remains an important taxonomic character.

Figure 1 Distribution of the species of Agave sect. Inermes and of A. ellemeetiana (sect. Choritepalae) in the biogeographic provinces of Mexico (Morrone 2017).
We aimed to describe and illustrate Agave rosalesii from western Oaxaca, Mexico, as a species new to science, placed in A. sect. Inermes as its twelfth species.
Materials and methods
As a result of fieldwork in the montane pine-oak forest of the Putla District in the Oaxaca state, Mexico, we documented the morphology ofAgave ellemeetianavar.subdentataTrel. The collections of flowers were made in December 2011 and of the rosette morphology and variability in April 2014. After an analysis of the morphological structures of these plants, we compared with similar Agave species treated in taxonomic reference works, particularly spineless or soft-spined ones (Gentry 1982, Vázquez-García et al. 2007a, b, Thiede 2001, 2020), as well as with digital images of specimens deposited in the herbaria ARIZ, CORU, MEXU, MO, US, XAL and ZSS.
We readily realized that the populations from the Putla District, Oaxaca, previously treated as Agave ellemeetiana K. Koch ssp. subdentata (Trel. in L. H. Bailey) Thiede, needed to be segregated from the A. ellemeetiana complex as a separate species given their differing flower structure.
We confirmed vegetative characters of the Agave populations from the Oaxaca Putla District with the vegetative type specimen at the SSZ herbarium and color plates available in the literature. We compared the new species with both A. ellemeetiana (sect. Choritepalae) and A. pedunculifera (sect. Inermes) (Table 1) and documented the habitat differences of both these species (Table 2).
Table 1 Morphological differences between Agave rosalesii, A. ellemeetiana and A. pedunculifera.
| A. pedunculifera | A. ellemeetiana s. s. | A. rosalesii | |
|---|---|---|---|
| Habit (stem) | Acaulescent | Acaulescent | Sometimes caulescent |
| Rosette (branching) Height (cm) Diam. (cm) |
Not offsetting 70.0-90.0 140.0-180.0 |
Surculose 35.0-50.0 70.0-100.0 |
Not offsetting 50.0-70.0 93.0-145.0 |
| Leaf wax bloom | Present | Absent | Present |
| Leaf shape | Mostly ovate-acuminate or lanceolate | Ovate to oblong | Broadly oblanceolate to rarely elliptic |
| Leaf size | 80.0-90.0 × 11.0-15.0 cm | 50.0-70.0 × 12.0-20.0 | 43.0-70.0 × 14.0-30.0 |
| Leaf margin | Narrowly lined with brown or white, with closely set denticles 0.5-2 mm long | Entire, sometimes finely serrulate towards the apex | Very minutely denticulate |
| Leaf length to width ratio | 5.1-5.3 | 4.4-4.6 | 2.6-3.2 |
| Terminal spine (mm) | Acicular, weak, 10 mm | Absent, but leaf tip shortly acuminate and slightly calloused | Shortly conic, 2-3 mm or inconspicuous, reddish |
| Inflorescence (m) | 2.0-3.0, erect to recurving | 3.0-4.5, erect | 3.2-3.5, erect |
| Flower pedicel length (mm) | 20.0-30.0 | 15.0-20.0 | 4.0-5.0 |
| Flower length (cm) | 3.7-5.2 | 2.8-4.0 | 3.3-3.5 |
| Corolla tube | Evident, 2.0-4.0 mm (as in A. sect. Inermes) | Inconspicuous, 1.0-2.0 mm (as in A. sect. Choritepalae) | Evident, 4.0-5.5 mm (as in A. sect. Inermes) |
| Flower color | Pale yellow | Pale greenish-yellow to white | Yellow to pale yellow |
| Tepal lobes (size in mm) | 12.0-18.0 × 5.0-6.0, erect | 13.0-15.0 × 5.0, incurved | 19.0-22.0 × 6.5-7.0, reflexed-rolled |
| Number of flowers per node | 4 | Usually 4 | 2 |
| Filament length (cm) | 4.0-5.5 | 5.0-6.0 | 6.0-6.5 |
| Anther length (mm) | 14.0-22.0 | 10.0-12.0 | 16.0-17.0 |
| Style length (mm) | 20.0-27.0, shorter than the filaments | 50.0-60.0, as long as the filaments | 40.0-53.0, shorter than the filaments |
| Fruit size (cm) | 1.5-2.0 × 0.7-1.0 | 1.3-1.5 × 1.0 | 2.0-2.3 × 1.1-1.6 |
Table 2 Habitat differences between Agave rosalesii, A. ellemeetiana, and A. pedunculifera
| A. pedunculifera | Agave ellemeetiana s. s. | A. rosalesii | |
|---|---|---|---|
| Latitudinal range (°N) Distribution |
19.747691 - 24.316667 Sinaloa, Nayarit, Jalisco |
18.031346-18.913517 Veracruz, Oaxaca (N) |
17.009966-17.157348 Oaxaca (W) |
| Altitude (m) | 1,500-2,000 | 400-1,600 | 2,400-2,600 |
| Vegetation type | Premontane pine-oak forest | Tropical subdeciduous forest to cloud forest | Montane pine-oak forest |
| Biogeographic province | Sierra Madre Occidental, Eje Neovolcánico Transversal | Sierra Madre del Sur (ESE) | Sierra Madre del Sur (SSE) |
| Physiography | Sierra de Tacuichamona Sierra de Tapalpa, Sierra de Tlacotepec | Sierra de Zongolica, Sierra Mazateca | Mixteca Alta |
| Hydrography (watersheds) | Río San Lorenzo, río Tapalpa, Bajo Balsas, Huetamo, La Salada | Papaloapan | Río Verde basin |
We used GeoCat (Bachman et al. 2011) online application to calculate the Extent of Occurrence (EOO) and Area of Occupancy (AOO) for their conservation assessment.
Results
Agave rosalesii A. Vázquez, Thiede, Etter & Kristen, sp. nov. (Figures 2-4). Type. Mexico, Oaxaca, distrito de Putla, municipio Santa Cruz Itundujia, paraje Los Monos, Col. El Chamizal, Buena Vista, bosque de encino-pino, creciendo sobre promontorios de roca con crasuláceas y Salvia, 16° 54' 24.4'' N, 97° 37' 22.4'' W, 2,549 m asl, 27 November 2013, Vicencio Aparicio-Barrios 278 w/ Kenia Velasco-Gutiérrez & Javier Juárez-Sierra (holotype: MEXU).
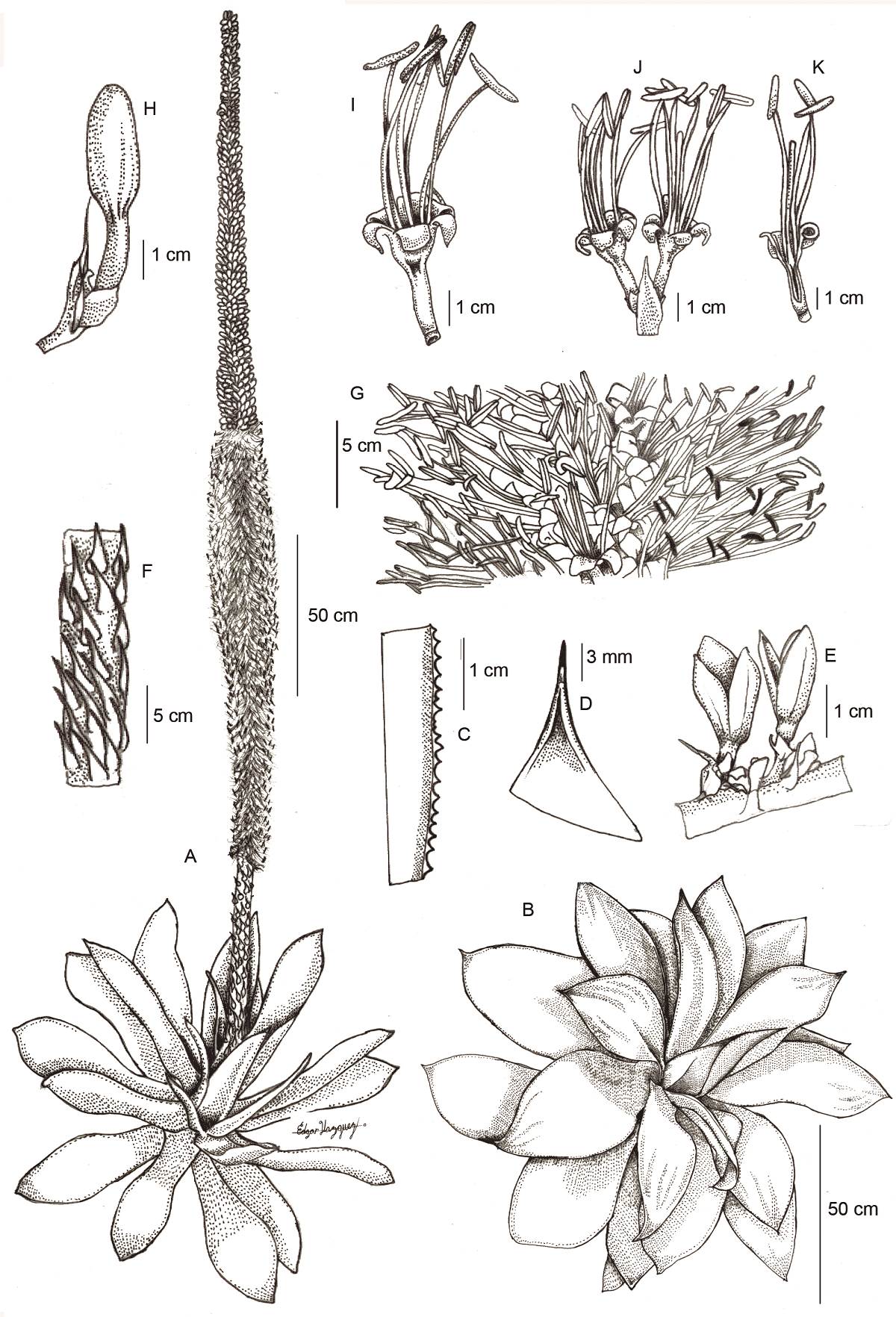
Figure 2 Agave rosalesii. A. Habit. B. Rosette. C. Leaf margin. D. Terminal spine. E. Capsules. F. Peduncle segment. G. Inflorescence segment. H. Flower bud. I. Fully developed flower. J. Geminate flowering node. K. Dissected flower. Mostly from photographs by Julia Etter & Martin Kristen, except E. from E. Solano w/ R. Ríos 1976 (MEXU). Illustration by Edgar Esau Vázquez-Verdejo.

Figure 3 Agave rosalesii. A. Geminate flowering node with outer and inner floral bracts (bracteoles). B. Dissected flower showing corolla tube and ovary. C. Inflorescence close up. D. Leaf showing a terminal spine. E. Habitat. F. Habit in full bloom. G. Rosettes. H. Leaf margin. Photographs by Julia Etter & Martin Kristen.

Figure 4 Agave rosalesii. A. Capsules, from E. Solano w/ R. Ríos 1976 (MEXU). B-C. Neotype of Agave ellemeetiana ssp. subdentata, J. Thiede s.n. (ZSS)
= Agave ellemeetiana var. subdentata Trel. in L. H. Bailey, Stand. Cycl. Hort. 1: 236 (1914)
≡ Agave ellemeetiana ssp. subdentata (Trel. in L. H. Bailey) Thiede, Bradleya 32: 157 (2014), syn. nov. - Type (neotype, designated by Thiede 2014: 157): ex cult., J. Thiede s.n., 8th October 2014; from living plants raised from seed collected in Mexico, Oaxaca, between Putla and Tlaxiaco, ca. 2400 m; J. Spath & K. Griffin s.n. (ZSS digital image seen!).
Diagnosis. Agave rosalesii shares with A. ellemeetiana the soft leaves, inflorescence size and shape, but differs from the latter in having rosettes non-offsetting (vs. surculose); leaf wax bloom present (vs. absent); leaves broadly oblanceolate to rarely elliptic (vs. ovate to oblong); leaf margin very minutely denticulate (vs. entire and sometimes finely serrulate towards the apex); leaf length to width ratio 2.6-3.2 (vs. 4.4-4.6); terminal spine 2.0-3.0 mm or inconspicuous (vs. absent); pedicels 4.0-5.0 mm long (vs. 15-20 mm); the number of flowers per node 2 (vs. usually 4); corolla tube 4.0-5.5 mm long (vs. 1-2 mm); tepal lobes reflexed-rolled, 19.0-22.0 × 6.5-7.0 mm (vs. incurved, 13.0-15.0 × 5.0 mm); anthers 16.0-17.0 mm long (vs. 10.0-12.0 mm); style 40.0-53.0 mm long (vs. 50.0-60.0 mm); and fruit size 2.0-2.3 × 1.1-1.6 cm (vs. 1.3-1.5 × 1.0 cm) (Table 1).
Agave rosalesii is also similar to Agave pedunculifera in having non-offsetting rosettes, a denticulate leaf margin, a distinct corolla tube, a similar tepal lobe size and anther length, but it differs from the latter in having leaves broadly oblanceolate to rarely elliptic vs. mostly ovate-acuminate or lanceolate; leaf size 43.0-60.0 × 14.0-30.0 cm (vs. 80.0-90.0 × 11.0-15.0 cm); leaf length to width ratio 2.6-3.2 (vs. 5.2); terminal spine 2.0-3.0 mm long (vs. 10.0 mm); inflorescence 3.2-3.5 m long (vs. 2.0-3.0 m); pedicels 4.0-5.0 mm long (vs. 20.0-30.0 mm); flower length 3.3-3.5 cm (vs. 3.7-5.2 cm); flower color yellow (vs. pale yellow); tepal lobes reflexed rolled, 19.0-22.0 × 6.5-7.0 mm (vs. erect, 12.0-18.0 × 5.0-6.0 mm); number of flowers per node 2 (vs. 4), filaments 60.0-65.0 mm long (vs. 40.0-55.0 mm); and style 40.0-53 mm long (vs. 20-27 mm) (Table 1).
Description. Plants perennial, sometimes caulescent and up to 100.0 cm long, stem usually absent or up to 50.0 cm long, rosettes 50.0-70.0 cm in height, 93.0-145.0 cm in diam., consisting of 14-28 leaves, not offsetting, leaves 43.0-70.0 × 14.0-30.0 cm, broadly oblanceolate or rarely elliptic, narrowed at the base, 10.0-15.0 cm wide, adaxially concave, glaucous-green to green-yellowish, with distinct leaf wax bloom, longitudinally wrinkled toward the apex, the apex arching outward, abaxially convex and pale bluish-green; leaf margin very minutely denticulate with soft herbaceous teeth, regularly spaced, whitish or reddish; terminal spine shortly conic, 2.0-3.0 mm or inconspicuous, reddish. Inflorescence spike-like, 320.0-350.0 × 24.0-26.0 cm, erect, densely flowered, the fertile part 250.0 cm long, peduncle 90.0-100.0 cm long, 4.5-5.0 cm in diam. at the base; upper peduncle bracts 6.0-7.0 × 1.0-1.2 cm at the base; outer floral bracts 1.4-1.5 cm long, inner flower bracts (bracteoles) 0.4-0.5 cm long; pedicels 4.0-5.0 mm long. Flowers geminate, 3.3-3.5 cm long, yellow; ovary 19.0-20.0 × 4.8-5.2 mm; style shorter than the filaments, 40.0-53.0 mm long, yellow; stigma capitate, yellow; corolla tube evident, 4.0-5.5 mm long; tepal lobes 19.0-22.0 × 6.5-7.0 mm, reflexed-rolled, yellow to pale yellow; filaments 6.0-6.5 cm long, creamy white to pale yellow; anthers 16.0-17.0 mm long, yellow. Fruits 2.0-2.3 × 1.1-1.6 cm, obovoid, fruit pedicels 6.0-8.0 mm; seeds unknown.
Distribution and ecology. Agave rosalesii is endemic to the Putla District in western Oaxaca, in the municipalities of Santa Cruz Itundujía (paraje Los Monos), Santa María Yucuhiti (Yosonicaje) and Putla Villa de Guerrero (Las Mesas Santo Domingo), inhabiting rocky outcrops of montane pine-oak forest at 2,400-2,600 m in elevation within the Río Verde basin. It grows with A. kavandivi, Echeveria sp., Furcraea sp., Pinguicula sp., Pinus sp., and Sprekelia sp. (Table 2, Figure 1).
Etymology. The species honors Carlos Santiago Rosales Martínez, a young enthusiastic agavologist, explorer, and horticulturist of succulents, who, in collaboration with the first author, unveiled that the Putla populations traditionally treated as belonging to A. sect. Choritepalae, actually belong to A. sect. Inermes due to the presence of a distinct corolla tube.
Conservation status.Agave rosalesii is locally abundant but confined to a few locations in a restricted area (EOO 45,941 Km2 and AOO of 12,000 km2). This species, meeting the A3(c) criterion, is critically endangered (CR). We expect a population reduction for the following decades, given current trends in climate change that will deteriorate habitat quality at the top of the mountain, depleting the only space left for survival. Additionally, corn crop expansion is also a threat since it restricts the agaves to the rocky outcrops, where corn plants could not succeed. The species has the potential as an ornamental for temperate regions.
Additional specimen examined (paratype). Mexico, Oaxaca, distrito de Putla, municipio de Putla Villa de Guerrero, Las Mesas Santo Domingo, 17° 09’ 21.86” N, 97° 50’ 37.37” W, 2,442 m asl, 13 April 2006 (fr), E. Solano w/ R. Ríos 1976 (MEXU).
Discussion
Agave populations from the Pacific slopes in the Putla District of western Oaxaca, formerly treated as Agave ellemeetiana ssp. subdentata (Thiede 2014) differ sufficiently in their flower and fruit morphology from disjunct populations of Agave ellemeetiana s. s., occurring in the Atlantic slopes of central Veracruz and northern Oaxaca (Cházaro-Basáñez et al. 2012, Thiede 2014). This evidence supports the recognition of A. ellemeetiana ssp. subdentata as a different species, here proposed as A. rosalesii sp. nov., and also places it in the A. sect. Inermes and not in A. sect. Choritepalae due to the presence of an evident corolla tube. Thus, withinAgave, this is the first record of an infraspecific taxon elevated as a species on a different section from the initial one, suggesting a revision of other poorly morphologically characterized taxa.
A phylogeny using 355 AFLP markers for 71 species of Agave s. s. also supports the separation of A. rosalesii, treated as A. aff. pedunculifera, from A. ellemeetiana s. s. (Gil-Vega et al. 2007). The Putla populations (A. rosalesii) cluster together with two species of A. sect. Inermes: A. pedunculifera and A. vilmoriniana. On the other hand, A. ellemeetiana was not clustered with any species. A cladogram based on nuclear DNA (ITS) shows A. bracteosa and A. ellemeetiana s. s., the two of A. sect. Choritepalae, as the most basal species in the genus (Jiménez-Barrón et al. 2020). This study shows that the Amolae and Choritepalae groups (now formally as A. sect. Inermes and A. sect. Choritepalae, respectively) are not monophyletic, suggesting the corolla tube is not a reliable predictor of phylogenetic relationships.
Ecological differences also separate these two taxa: Agave rosalesii is a montane species thriving from 2,400 to 2,600 m, while A. ellemeetiana s. s. grows in lowlands or mid-elevations, from 400 to 1,600 m (Cházaro-Basáñez et al. 2012) (Table 2). Among the species of A. sect. Inermes, only two grow at elevations above 2,400 m: A. rosalesii in the Mixteca Alta of western Oaxaca and A. manantlanicola in the Sierra de Manantlán of Western Mexico; the former with soft broad leaves, spineless or soft-spined and subdentate margin, and the latter with stiff leaves, long firm-spine and corneous toothless margin (Cuevas-Guzmán et al. 2012, Thiede 2014). Also, the latitudinal range of A. rosalesii, growing at about 17° N, is further south than that of A. ellemeetiana s. s., growing from 18-19° N. Actually, A. rosalesii is the second southernmost species of A. sect. Inermes, and also the second recorded for Oaxaca (Figure 1). The two species inhabit different habitats: A. rosalesii grows in a montane pine-oak forest in the Pacific slopes within the Río Verde watershed with a mean annual rainfall of 1,800 mm, while A. ellemeetiana s. s. inhabits tropical deciduous to cloud forest in the Atlantic slopes of the Papaloapan watershed with a mean annual rainfall of 2,800 mm (Table 2) (Thiede 2014).
A conservation strategy needs to be designed, we strongly recommend setting aside a natural protected area to ensure the survival of the critically endangered species, Agave rosalesii, and all together protect the rich and the endemic flora of the montane pine-oak forest of the Mixteca Region, an area of numerous recent discoveries, such as the corncob flower, Magnolia mixteca (Vázquez-García et al. 2021), the cycad Dioon planifolium Salas-Mor., Chemnick & T.J. Greg. (Salas-Morales et al. 2016), Salvia robertoana Mart.Gord. & Fragoso (Martínez-Gordillo et al. 2016) and Salvia tilantongensis J.G. González & R. Aguilar-Santelises (González-Gallegos & Aguilar-Santelises 2014).
Key to species of A. sect. Inermes similar to A. rosalesii
1. Plants surculose; mature rosettes on stems longer than 50.0 cm.
2. Stems 20.0-25.0 cm diam.; leaf margin corneous, continuous and entire; terminal spine 30.0-45.0 mm long; tepals reddish outside .................................................................................... A. manantlanicola
2. Stems less than 15.0 cm diam.; leaf margin smooth or serrulate; terminal spine absent; tepals yellowish green ........................................................................................................................... A. attenuata
1. Plants non-surculose; mature rosettes acaulescent or on stems less than 30.0 cm long.
3. Terminal spine 20.0 mm long or less.
4. Leaf margin closely denticulate; corolla tube 2.0-6.0 mm long
5. Leaves 80.0-90.0 × 11.0-15.0 cm, terminal spine 10.0 mm long, pedicels 20.0-30.0 mm long, tepal lobes 12.0-18.0 × 5.0-6.0 mm long and pale yellow, filaments 40.0-55.0 mm long, styles 20.0-27.0 mm long ....................................................................................................................... A. pedunculifera
5. Leaves 43.0-60.0 × 14.0-30.0 mm, terminal spine 2.0-3.0 mm long, pedicels 4.0-5.0 mm long, tepal lobes 19.0-22.0 × 6.5-7.0 mm long and yellow, filaments 60.0-65.0 mm long, styles 40.0-53.0 mm long ..................................................................................................................................... A. rosalesii
4. Leaf margin entire; corolla tube 11.0-12.0 mm long ...................................................................................................................................... A. gilbertii
3. Terminal spine 30.0-65.0 mm long.
6. Leaf margin entire; flowers 20.0-30.0 mm long; tepals ivory-white .................................................................................................................................... A. chazaroi
6. Leaf margin closely dentate; flowers 52.0-94.0 mm long, tepals greenish-yellow ...... A. vazquezgarciae

