











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
The Late Cretaceous is one of the most significant periods in the geological history of the Earth planet, considering its end with the famous mass extinction of many genera and species of animals [ Alvarez et al. (1980), Raup et al. (1982), Molina et al. (1996), Bambach et al. (2006), Schulte et al. (2010), Keller et al. (2010), Archibald et al. (2010), Longrich et al. (2011), Mc Ghee et al. (2013), Brusatte et al. (2015), James et al. (2016)]. The Aurès Basin in the Eastern Saharan Atlas is one of the most intriguing regions of northeastern Algeria. It includes major structural and tectonic events that have affected the region. As a result of this tectonic activity, the stratigraphic sequence records a greater complexity of facies changes and fluctuations in microfaunal contents (foraminifera in particular) that also reveal changes in the depositional environment.
In this work, which is based on data obtained in 2016 (Benmansour, 2016) and already partly published (Benmansour et al., 2017) - where the authors explain how they defined the boundaries of the stages and sub-stages- we focused on the genus Laffitteina in Maastrichtian-Thanetian deposits. This genus was established in 1945 by Marie based on material from the Montian of France. The test of this genus is lenticular with a diameter of 3 mm and a thickness of 1 mm (Blanc, 1975). The members of this taxa have in common a single foramen, slit-shaped, straight, and areal, extending from a ventral interiomarginal position in an oblique direction into the dorsal parts of the septal face. They exhibit no foliar chamberlets but a plate of which the nature and its relation to the areal foramen is unclear (Hottinger, 2014).
Laffitteina is one of the few groups of larger, complex foraminifera surviving the Cretaceous-Cenozoic boundary event in shallow water (Billman et al., 1980). It is widely present in the Maastrichtian from the Pyrenean Basin to the Middle East platforms (Consorti et al., 2018). It occurs also in the Paleocene of the Middle East (Rahaghi, 1992; İnan et al., 2004) where its stratigraphic distribution through the Cretaceous and Paleocene carbonates is practically continuous (Hottinger 2014, in Consorti et al., 2018).
According to Billman et al. (1980) and Hottinger (2014), Laffitteina thrive in meso- to eutrophic environments. The latest Maastrichtian-Danian low-stand phase [(Hallam and Wignall, 1999; Alegret et al., 2001; Habib and Saeedi, 2007; Haq, 2014)] may have increased the tropism in shallow seas through the establishment of “pools” restricted in water circulation on shelves. This would provide environment for some resistant foraminifera represented by Laffitteina, Elazigina, and probably Palaeoelphidium (Consorti et al., 2018). Belkhoudja et al., (2004) are also mentioned that the dominance of the Laffıtteina implies the installation of lagoon setting with decreased salinity (up to 30 ‰).
In Algeria, the genus Laffitteina was first reported by Drooger (1952) by only two species: L. bibensis and L. conica. Subsequently, six species were recognized at least in the outcrop of El Kantara, western Aurès (Belkhodja and Bignot, 2004), namely: L. boluensis (Dizer, 1957); L. conica (Drooger, 1952); L. mengaudi (Astre, 1923); L. cf. monodi (Marie, 1946); L. oeztuerki (Inan, 1995) and L. bibensis (Marie, 1946).
The purpose of this paper is to describe the microfacies and assemblages of Laffitteina from the shallow marine carbonate of the Maastrichtian-Thanetian of the Aurès Basin and to establish deposits conditions at the K-Pg boundary.
2. Geological setting
The Upper Cretaceous - Paleocene sedimentary rocks are well exposed in the Aurès basin in the eastern part of the Saharan Atlas (northeastern Algeria) (Figure 1A). The Aurès has previously been mapped by Laffitte (1939), stratigraphicaly, sedimentologically and structurally studied by Dubourdieu (1956), Bertraneu (1955), Cruys (1955), Emberger (1960), Guiraud (1973, 1974, 1975), Vila (1980), Bureau (1986), Kazi Tani (1986), Aissaoui (1985), Ghandriche (1991), Addoum (1995) and Herkat (1999).
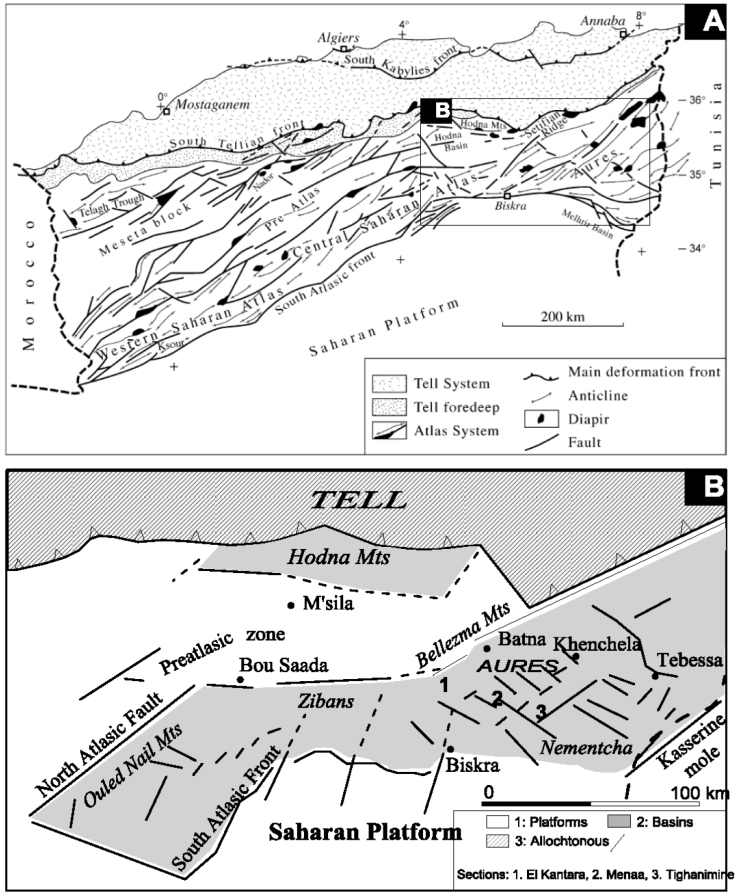
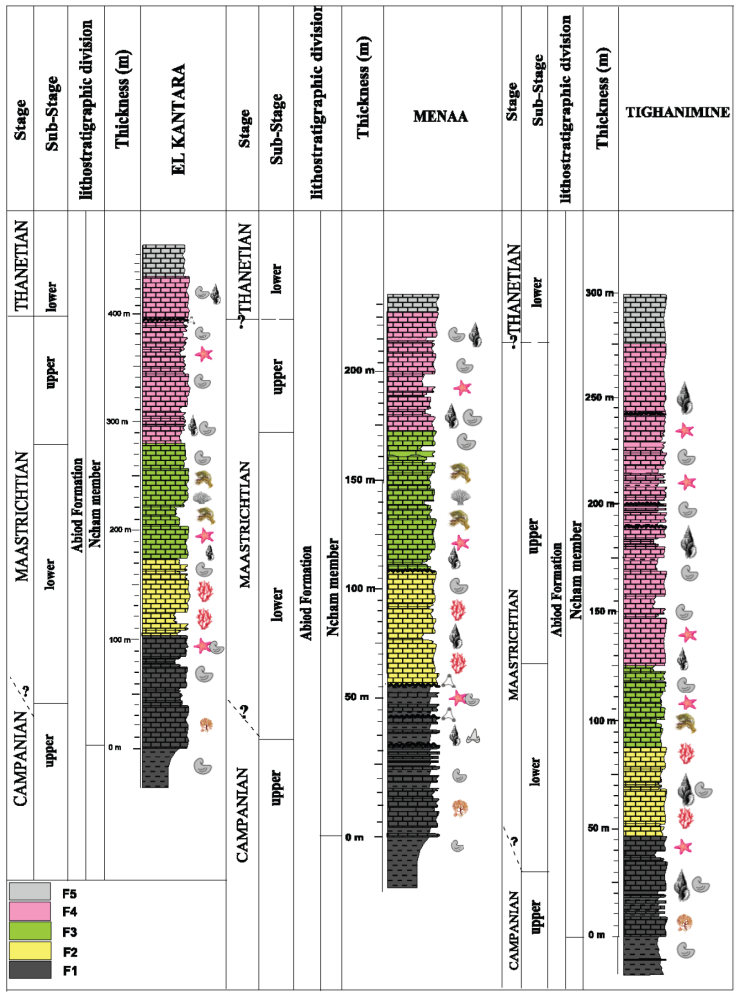
Figure 1 A. Main structural domains of northern Algeria and location of the studied area in the eastern part (modified from Michard et al., 2008). B. Paleoenvironments of the Eastern Atlasic Domain (Herkat & Guiraud, 2006) and location of the sections, from West to East: El Kantara, Menaa, and Tighanimine.
The succession, named by Benmansour et al. (2017) Ncham Member, belongs to be upper Campanian-Maastrichtian and Thanetian.
The Aurès Basin is characterized by a system of tilted blocks bordered by NW-SE to WNW-ESE-trending faults Guiraud (1973, 1990), Bellion et al. (1973), Vila (1980), Bureau (1986) et Kazi Tani (1986). Otherwise, NE-SW faults located within the basin are characterized by transtentional movements. During the late Maastrichtian-Paleocene, the structural evolution in the Aurès Basin led to the development of slumps with slip parallel to the dip of the structure limbs, as seen in the upper Maastrichtian formations of the Tebessa Mountains (Herkat, 1999). Also, progressive unconformities of Paleogene on top of the Maastrichtian are developed on the southern flank of the Djebel El Azreg anticline. These features suggest the onset of folding in the Aurès’ anticlines at the end of the Cretaceous (Herkat et al., 2006). A Palaeocene sedimentary break is observed at Djebel Azreg, in the central zone of the Aurès Massif, that the Eocene sediments unconformably overlying the Maastrichtian, whereas farther west this stage is recognized at Djebel Metlili, in the northern part of the Ziban range (Belkhodja and Bignot, 2004).
3. Materials and Methods
In this study, we use the same lithostratigraphic nomenclature adopted by Benmansour (2016) and Benmansour et al., (2017) for the upper portion of the Upper Cretaceous-Lower Paleogene deposits of the Aurès Basin that constitute the Abiod Formation. The Ncham Member, representing the Maastrichtian-lower Thanetian limestones rocks (the upper part of the Abiod Formation), are sampled from three localities (Figure 1B) in the western Aurès (El Kantara section: GPS: 35°13’57” N and 5°42’6,93” E, with a thickness of 450 m), and the central Aurès, on both sides of Djebel El Azereg (Menaa GPS : 35°11’08. 44” N and 5°59’0 42,00” E, with a thickness of 200 m and Tighanimine: GPS : 35°9’30,19” N and 6°14’29,03” E, with a thickness of 300 m). A total of 232 thin sections were prepared from samples of hard limestones that collected from the localities described above using a monocular polarizing microscope, in order to describe microfacies and study the populations of Laffitteina in random sections. The number and kinds of samples depend on the geological settings (outcrop conditions, sampling units indicated by types and thickness of bedding and bed by bed sampling was conducted several times, if lithological changes were observed within the bed). Carbonate rocks in this study were classified following Dunham (1962), Folk (1959), and Embry and Klovan (1971) for reefal rocks texture. The palaeoenvironmental reconstruction is based on Wilson (1975); Tucker and Wright (1990); Wright (1992); Scholle and Ulmer-Scholle (2003), and Flügel (2004). All the thin sections containing the Laffitteina species figured in this paper using a Sony Cyber-shot DSC-W670 16.1MP 1/2.3” CCD Compact Camera are deposited at the collection of Paleontology Laboratory, of the Geological Department, Batna 2 (Mostefa Ben Boulaïd) University, Algeria, under the collection label “K” for El kantara section, “M” for Menaa section and “T” for Tighanimine section, respectively.
4. Stratigraphic Setting
In northeastern Algeria, the Abiod Formation marks its location in the upper portion of the Late Cretaceous-Early Paleogene very clearly by its effect on the topography, as mentioned in north-central Tunisia by Robszynski et al., (2010): a double line of low hills separated by a slight median depression. The Upper Cretaceous-Paleocene of the Aurès is organized in the landscape as double hills, separated by a middle part in a slight depression. This morphology corresponds to the tripartite composition of the Upper Cretaceous-Lower Paleocene with a lower member of alternances of marl-limestone, a middle member constituted mainly by marls and an upper member of massive and often chalky limestones, with a bright white patina (equivalence of Ncham Member in Kalaat Senan). The regularity of this trilogy is modified in some places by reductions, or even lacunae, visibly affecting the upper member. In this study, there is a clear reduction in thickness of the Ncham Member in Menaa section, that explained by its location at the apex of tilted blocks and therefore the reduction in the space available for sedimentation.
This study is based on Ncham Member from three localities as mentioned above (Figure 1B): El Kantara section, Menaa and Tighanimine section. This member is of Late Cretaceous-Early Paleogene age. As cited in 2016 (Benmansour, 2016) and Benmansour et al. (2017), the Campanian is identified by Globotruncanita elevata, Globotruncana ventricosa, Radotruncana calcarata, Globotruncana falsostuarti and by the Ammonite: Nostoceras (Bostrychoceras) polyplocum (ROEMER, 1841). The Campanian-Maastrichtian boundary could not be specified with the foraminifera. Nevertheless, it is located between the disappearances of Globotruncana falsostuarti and Nostoceras (Bostrychoceras) polyplocum. The Maastrichtian is highlighted in most sections by the existence of planktonic foraminifera Rugoglobigerina macrocephala and Rugoglobigerina hexacamerata. The lower Maastrichtian-upper Maastrichtian boundary was traced by the first appearance of the stratigraphic species Omphalocyclus macroporus which corresponds to the upper Maastrichtian. The Cretaceous-Paleogene boundary was identified by the first occurrence of Laffitteina erki which marks the Thanetian age.
In the El kantara section, the Ncham Member consists of about 450 m of massive limestones, which form the famous gorges of El Kantara. The member begins with bioclastic limestones very rich in bryozoans, echinoids, and Inoceramus. In the topmost of the member, bioclastic limestone beds become more common and are overlay by limestones with calcareous algae and Laffitteina. It is bounded by an erosional surface of indurated bioclastic limestones containing thalassinoides. Placed in the paleogeographic sketches proposed by the authors and more particularly on those established by Vila (1980) and Tawadros (2001), the El Kantara deposit appears to be located at the beginning of the trans-Saharan sea, at a short distance from the emerged lands of the Paleomaghreb. The Cretaceous-Thanetian period ends with a shallow marine sedimentation, very changeable, passing from infralittoral to mediolittoral and then to supralittoral (Belkhoudja, 2004). The return of the sea begins in the upper Thanetian by the installation of a series of lakes where the Red Marls are deposited above the Ncham Member. The constant presence of Microcodium in these marls is an indication of the proximity of emerging limestone outcrops in the process of karstification whose erosion brings into the gulf puffs of coarse terrigenous contributions: quartz and especially limestone micro-breccias, remanified foraminifera, articles of Microcodium (Belkhoudja, 2004).
In the Menaa section, the Ncham Member consists of about 200 m of massive limestones. It begins with a marl-limestone unit, in cm-to dm-thick beds, containing abundant bivalves, gastropods, and bryozoans. This unit ends with intensively bioturbated limestone beds with traces of thalassinoides. The upper part of the member consists of about 100 m of monotonous bioclastic limestone containing abundant bivalves, echinoids, gastropods, and bryozoans. At the top, calcareous algal beds have been recorded.
In the Tighanimine section, the Ncham Member consists of about 300 m of massive bioclastic limestones. It constitutes an easily identifiable landmark in the topography of the Aurès. It presents two aspects, successively from the bottom to the top: 1- several levels of lime-stone appear in centimetric beds, sometimes bioclastic or with lumachellic aspect; with a progressive thickening towards the top, ending with an emersion surface, 2- the lime-stone becomes massive containing abundant accumulations of bivalves, echinoids, gastropods, bryozoans, and red algae.
5. Results
5.1. Systematic Paleontology
Species of Laffitteina from the Maastrichtian-Thanetian shallow-water carbonates of the Aurès Basin (northeastern Algeria) are described herein. The vertical distribution of species is shown in Figures 4, 5 and 6.
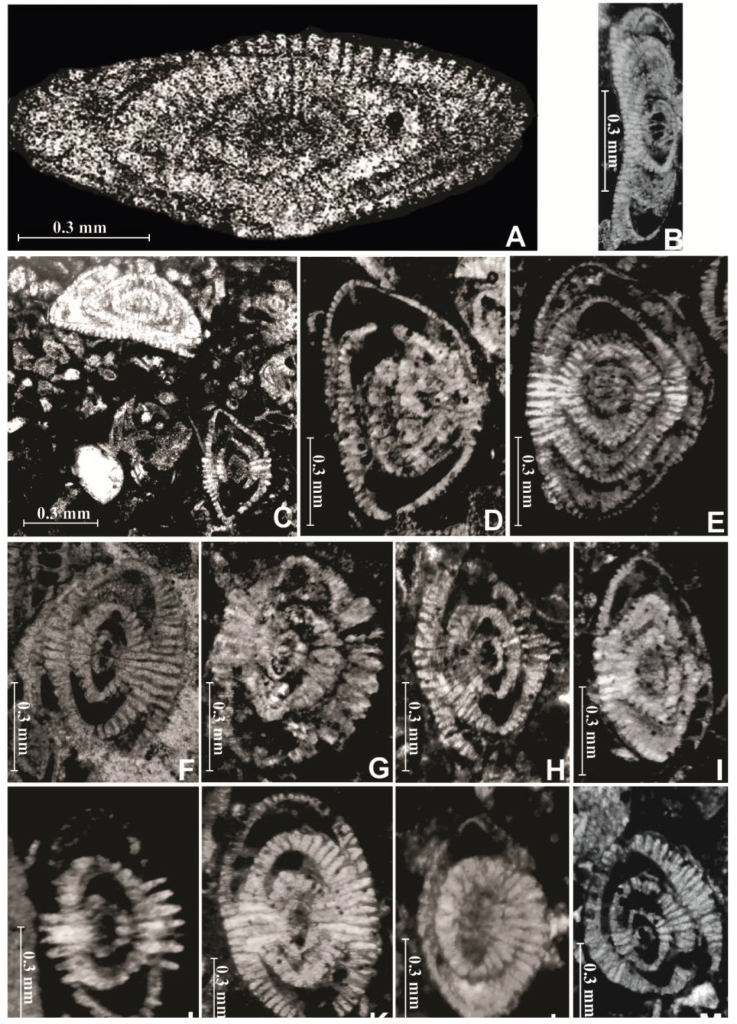
Figure 2 Laffitteina foraminifera in thin sections in the limestone beds of the Ncham Member. A, Laffitteina mengaudi. Axial section from El Kantara section. Sample K 71. B, Laffitteina aff. mengaudi. Axial section from El Kantara section. Sample K64. C-E, Laffitteina conica. Axial sections from El Kantara section. Samples K 44, K 54, K 74 and K 80 respectively. F-I, Laffitteina oeztuerki. Axial sections from El Kantara section. Samples K 23, K 65, K 73, K 82 respectively. J-M, Laffitteina marsicana, J&K. Axial sections from El Kantara section. Samples K 64 and K 77; L, Axial section from Menaa section. Sample M 71; M. Axial section from Menaa section. Sample T 22.
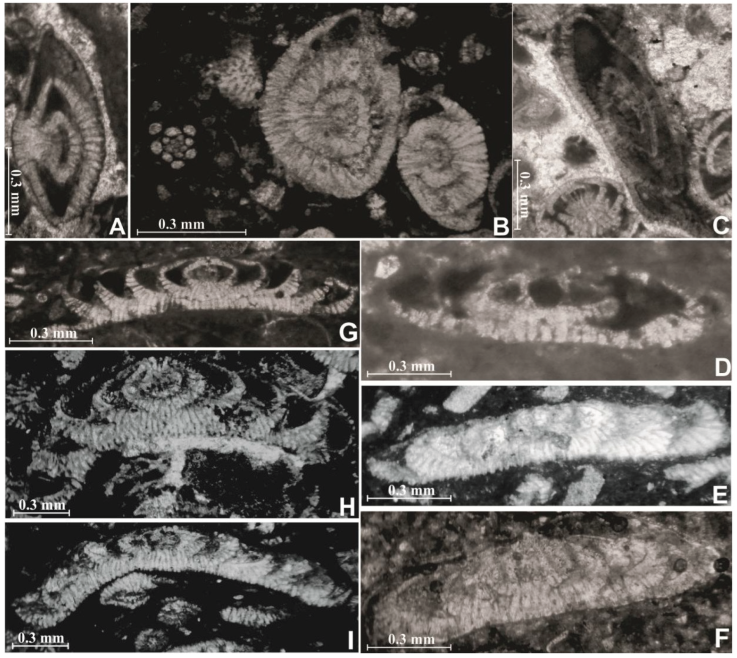
Figure 3 Laffitteina foraminifera in thin sections in the limestone beds of the Ncham Member. A-B, Laffitteina boluensis. Axial sections from El Kantara section. Samples K 39 and K 54. C-D, Laffitteina monodi. Axial sections from El Kantara section. Samples K 59 and K 82. E-G, Laffitteina bibensis, Axial sections from El Kantara section. Samples K 39, K 74, K88 respectively. H-I, Laffitteina erki, Axial sections from El Kantara section. Samples K 83 and K 89. I, Laffitteina aff. mengaudi. Axial sections from El Kantara section. Sample K64.
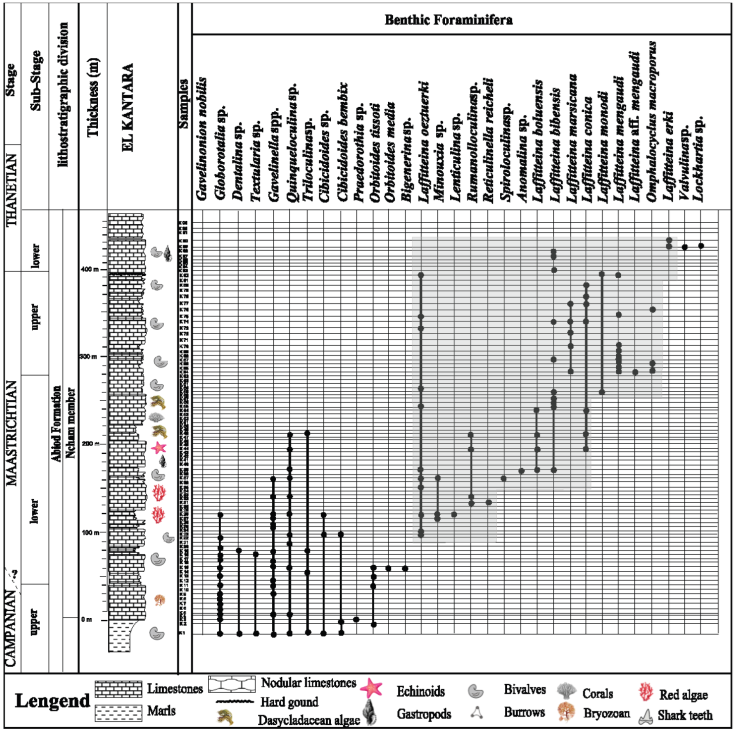
Figure 4 Vertical distribution of benthic foraminifera in El Kantara section, according to the order of the first appearance (Note: the legend in this figure is generalized also for the Figure 5 and Figure 6).
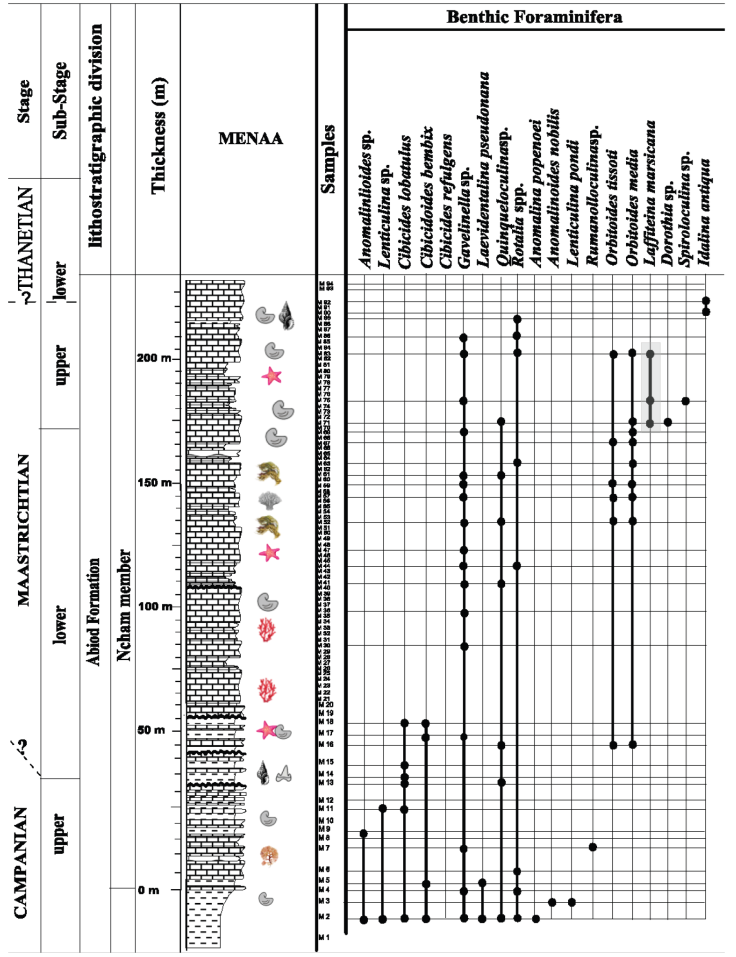
Figure 5 Vertical distribution of benthic foraminifera in Menaa section, according to the order of the first appearance.

Figure 6 Vertical distribution of benthic foraminifera in Tighanimine section, according to the order of the first appearance.
Order Foraminiferida Eichwald, 1930
Suborder Rotaliina Delage and Hérouard, 1896
Superfamily Rotaliacea Ehrenberg 1839
Family Rotaliidae Ehrenberg 1839
Subfamily Laffitteininae Hottinger, 2014
Genus Laffitteina Marie 1946
Type species.
Laffitteina bibensis Marie, 1946
Laffitteina mengaudi (Astre, 1923)
1923 Nummulites mengaudi Astre, p. 360-368, Pl.12.
1945 Laffitteina bibensis n. gen. n.sp., Marie, p. 419-431, figure 17-29
1945 Laffitteina monodi Marie, p. 419-431, figure 17-29.
1952 Laffitteina bibensis Marie, Drooger, p. 99-100,pl. 16, figure 9a, c, 11-15.
1957 Laffitteina cf. bibensis marie, Dizer, p. 38-41, pl. 1,2.
1957 Laffitteina boluensis s.sp. Dizer, p. 38-41, pl. 2.
1965 Laffitteina marsicana s.sp., Farinacci, p. 1245-1257, pl. 115-116.
1975 Laffitteina mengaudi (Astre), Blanc & Colin, p. 109-131, pl. 23-26.
1975 Laffitteina mengaudi (Astre), Blanc, p. 62, pl. 1-2.
1986 Laffitteina mengaudi (Astre), Sirel, p. 126, pl. 3.
1988 Laffitteina mengaudi (Astre), Radoicic, p. 22-24, pl. 4.
1998 Laffitteina mengaudi (Astre), Sirel, p. 88, pl. 46, figures 1-23; non pl. 47, figures 1-22.
Description. The test is inflated lenticular with broadly rounded periphery. In the axial section, umbilical and ventral sides are flattened or slightly convex. It has slightly trochospiral to nearly planispiral coiling.
Remarks. The test shape and the coiling type show similar characteristics to those of L. mengaudi illustrated by Marie from France (1946) and from Turkey (Sirel, 1996-1998).
Distribution. In this study, this specimen has been recorded from the late Maastrichtian of El Kantara section, at the level of samples K64-K70, K75, and K80. It occurs in the Maastrichtian-Paleocene limestones of Turkey (Sirel, 1996-1998) and in the late Maastrichtian of France (Marie, 1946).
Laffitteina aff. mengaudi (Astre, 1923)
Description. The test is thin lenticular with a flattened shape and rounded periphery. It has a flat ventral side, while the umbilical side is slightly convex. The coiling is slightly trochospiral.
Remarks. This specimen is closely similar to that recorded from Turkey and described by Sirel (1996). It shows close similarity with Laffitteina mengaudi in having rounded periphery and trochospiral coil, but it differs from L. mengaudi in having thinner lenticular shell.
Distribution. It was recorded only in sample K 67, from the late Maastrichtian of El Kantara section. It was found in the Maastrichtian-Paleocene of Turkey (Sirel, 1996).
Laffitteina conica Drooger, 1952
1952 Laffitteina conica Drooger, p. 100, Plate 16, figure 10a-c, 16,17.
1996 Laffitteina conica Drooger, Sirel, p.20, Plate V, figure 1-12; Plate, Plate VII, figure 1-17.
1998 Laffitteina aff. mengaudi (Astre)-Sirel, pars, p. 90, pl. 48, figure16.
Description. The shell is conical, with a narrowly rounded margin and apex. The axial section is asymmetric, planispiral involute, and has a triangular outline. It has a flattened umbilical area, where the pillars are strongly thickened. The spiral side is convex to strongly convex.
Remarks. The asymmetrical axial section and the thickness of the pillars on the umbilical side are similar to those of L. conica described by Marie (1946) and Sirel (1996-1998).
Distribution. In this study, this species was found only in the upper Maastrichtian of El Kantara section, in samples K44, K48, K 54, K 74, K 77, and K 80. It is very rare in Turkey; it has been found in the Maastrichtian limestones of Bozandere place, Ilgaz mountains, north of Çankiri and in central Turkey (Sirel, 1996-1998). It is associated in this locality with Laffitteina aff. mengaudi and Laffitteina mengaudi.
Laffitteina oeztuerkiInan, 1995
1995 Laffitteina oeztuerki n. sp.: Inan, p. 111, pl. I, figures 5-11.
1998 Laffitteina koyulhisarica n. sp.: Sirel, p. 92, pl. 52, figures 1-22.
Description. The test is lenticular biconvex, wider in the central part. It has an almost bilateral symmetric shape with a subacute periphery. The plugs are prominent on both sides.
Remarks. This species differs from other species of Laffitteina in having a lenticular, biconvex and nearly bilateral symmetric. This species is comparable to those described by Inan (2004).
Distribution. It occurs in the upper Maastrichtian of Turkey (Meriç and Inan, 1995; Inan, 2004). In this study, it was recorded in the upper Maastrichtian of El Kantara section, in samples: K 22, K 23, K 28, K 35, K 37, K 39, K 55, K 60, K 73, K 75, and K82.
Laffitteina marsicana Farinacci, 1965
1965 Laffitteina marsicana Farinacci, p? Pl ?, figure 8.2A-D.
1996 Laffitteina marsicana Farinacci: Inan, pl. I, figures 3-6.
Description. It has a thick lenticular test with a rounded periphery. The umbilical side is more convex than the ventral side, which may, however, have a slight umbonal swelling. It has asymmetrical planispiral coiling.
Remarks. This species is distinguised from the other species of Laffitteina by its plano-biconvex and lenticular test.
Distribution. In the study area, it occurs in the early upper Maastrichtian of El kantara section, in samples K 64, K 70, K 72, K 74, and K 77; in the Menaa section, in samples: M 71, M 75 83 and in the Tighanimine section, in samples: T 22, T 24 and T 25. It is found also in the early upper Maastrichtian of Turkey (Inan, 1996).
Laffitteina boluensisDizer, 1957
1957 Laffitteina boluensis Dizer, n. sp, pl. 2, figures 3-8
Description. The test is lenticular, biconcave with a planispiral coiling and an almost symmetrical shape. The margins are clearly blunt. It has thickened pillars, only on the umbilical side.
Remarks. This species shows similarities with L. monodi, but differs from the latter by its more compact septa and less loose equatorial section.
Distribution. It occurs in the late lower Maastrichtian of El Kantara section, in samples K 39, K 44, K K 48, and K 54. It was found in the middle Maastrichtian to the upper Danian of Turkey (Dizer, 1957).
Laffitteina monodiMarie, 1946
1998 Laffitteina aff. mengaudi (Astre)-Sirel, pars, p. 90, Pl.48, figure16.
Description. The test is lenticular with an irregular shape and a rounded periphery. The shell is compressed, with flattened umbonal areas on both sides. It has a semi-evolute coiling. Laffitteina monodi is distinguished from Laffitteina mengaudi by its more compressed shell.
Remarks. The present specimen differs from L. bibensis by its general irregularity and by the decrease of its spiral side. It is closely similar to L. monodi illustrated by Marie (1946).
Distribution. In this study, this species was found only in the upper Maastrichtian of El Kantara section, in samples K 59 and K 82. It also present in the Montian of Mauritania (Marie, 1946).
Laffitteina bibensisMarie, 1946.
1923 Nummulites mengaudi n. sp.: Astre, p. 360, pl. 12
1946 Laffitteina bibensis n. sp.: Marie, p. 419, pl. V, figures 1-3, 14-16.
1973 Laffitteina mengaudi (Astre): Blanc, p. 133, pls. 17, 18.
1975 Laffitteina mengaudi (Astre): Blanc, p. 61, pls. 1, 2.
1987 Laffitteina bibensis Marie-Loeblich and Tappan, p. 661, pl. 759, figures 1-2, 9.
1996 Laffitteina mengaudi (Astre): Sirel, pl. I, figures 1-22, pl. II, figures 1-22, pl. III, figures 1-11.
1998 Laffitteina mengaudi (Astre): Sirel, p. 88, pl. 46, figures 1-23, pl. 47, figures 1-22.
Description. The test is lenticular, almost planoconvex, or slightly biconvex. The chambers are hugging in asymmetrical chevrons. It has a flattened ventral side, while the umbonal side is flat (Figure 3F and 3G) or gently convex (Figure 3E),
Remarks. The specimen corresponds perfectly to the French specimens described and illustrated by Marie (1946; p 430, figures 14-16). But it shows a distinct pattern in the convex umbonal side.
Distribution: In this study, It was recorded in the upper Maastrichtian to lower Thanetian of El Kantara section, in samples K 39, K 55, K 56, K 57, K 59, K67, K 74, K 83, K 87, and K 88. It is present in the lower Danian of northwestern Europe (Bignot, 1987), in the Danian of Italy (Beckmann et al., 1982), in Maastrichtian of Tunisia (Bismuth et al., 1967), in the Paleocene of Libya (Berggren, 1974), in Maroc (Rahhali, 1970), in the Paleocene of Iran (Rahaghi, 1976, Bignot and Neumann, 1991), upper Maastrichtian-Danian of southwestern Slovenia (IGCP 286, 1991), Danian of Turkey (Fleury et al., 1985; Serra-Kiel et al., 1998), in the Adriatic (Drobne et al., 1995), Slandian of southwestern Spain (Serra-Kiel et al., 1996 and 1998).
Laffitteina erki (Sirel, 1969)
1957 Laffitteina cf. bibensis Marie, Dizer, pl. 1, figures 1-6, pl. 2, figures 1 and 2.
1969 Orduina erki n. sp.: Sirel, pl. 2, figures 1-5. 1969 Orduina erki var. conica n. var.: Sirel, pl. 3, figures 1-5.
1987 Orduina erki Sirel: Inan, pl. 8, figures 1-7. 1987 Orduina aff. erki conica Sirel: Inan, pl. 8, figure 8.
1994 Laffitteina erki (Sirel): Sirel, p. 47, pl. I, pl. II.
1995 Laffitteina erki (Sirel): Inan, pl. 2, figures 1-5.
1998 Laffitteina erki (Sirel): Sirel, p. 91, pl. 51, figures 1-8.
Description. The test is large and conical-shaped. It has a broadly rounded margin. The umbilical side is strongly concave, the spiral side is convex. Pillars Thickened only on the umbilical side.
Remarks. The specimen resembles L. erki described and illustrated by Sirel (1994 and 1996). But, it differs in being more elongated and arched.
Distribution. It occurs in the Thanetian lime-stones of Turkey (Sirel, 1996), and the lower Thanetian of El kantara section, in samples: K 83, K86, K 87, and K 89.
5.2. Microfacies description
Figures 4, 5 and 6 represent the distribution of benthic foraminiferal associations found in the study area. Figure 7 presents the results of the petrographic analysis and the abundant components from the Ncham Member. Five main microfacies were distinguished in the studied sections, where their vertical distribution was highlighted in Figure 8:
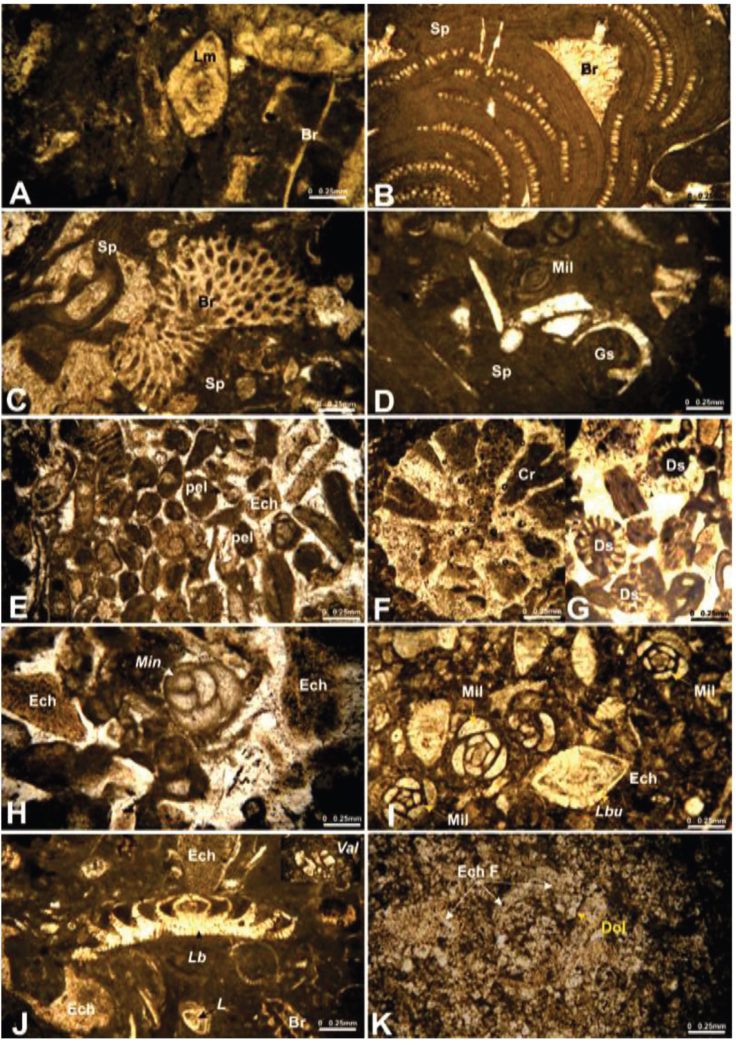
Figure 7 Thin sections of microfacies. A: F1: Bioclastic wackestone to packstone; B-D: F2. Algal bindstone Algal bindstone ; E-H: F3. Peloidal packstone to grainstone; I-J: F4. Foraminiferal wackestone to packstone; K. F5. Dolomitic mudstone. Abbreviations: Lm. Laffitteina marsicana; Br. Bryozoa; Sp. Sporolithon; Mil. Miliolids; Gs. Gastropods; pel. Peloids; Ech. Echinoids; Cr. Corals; Ds Dasycladacean algae; Min. Minouxia; Lbu. Laffitteina boluensis; Val. Valvulina; Lb. Laffitteina bibensis; L. Laffitteina; Dol. Dolomites; EchF. Echinoid phantoms.
Microfacies (F1): Bioclastic wackestone to packstone - Laffitteina, as well as miliolids, are rare constituents in this microfacies. Major grain types include bioclasts, which are composed of echinoids and bryozoans (Figure 7A). These allochems are bound by a micritic matrix.
Microfacies (F2): Algal bindstone - This microfacies includes scattered bioclasts of bryozoans, gastropods, ostracods, and some benthic foraminifera (particularly Laffitteina), which are encrusted and bound by red algae (Sporolithon) (Figure 7B-7D). Microfacies (F3): Peloidal packstone to grainstone Major grain types include peloids, bioclasts, and benthic foraminifera. Bioclasts are composed of dasycladacean algae, corals, bivalves, gastropods, and echinoderms, which exceed 2 mm in diameter. Benthic foraminifera are present notably by Laffitteina oeztuerki, Minouxia sp., Lenticulina sp., Rumanolloculina sp., and Reticulinella reicheli. These grains are cemented by a sparitic calcite cement (Figure 7E-7H).
Microfacies (F4): Foraminiferal wackestone to packstone - The allochems are mainly composed of Laffitteina (L. bibensis, L. oeztuerki, L. marsicana, L. conica, L. monodi, L. mengaudi, and L. aff. mengaudi,...) and miliolids. There are also very low proportions of scattered Anomalina, echinoids fagments, and bryozaons in this microfacies. The bonded material consists of micrite (Figure 7I-7J).
Microfacies (F5): Dolomitic mudstone - The Laffitteina limestones that constitute the micro-facies F4 are terminated by dolmitized lime-stone of microfacies F5 microfacies. It consists of small homogeneous rhombohedral crystals of dolomite, with a size of about 100 µm associated with chrinoids phantoms (Figure 7K). These grains are cemented with a clear micrite.
6. Discussion
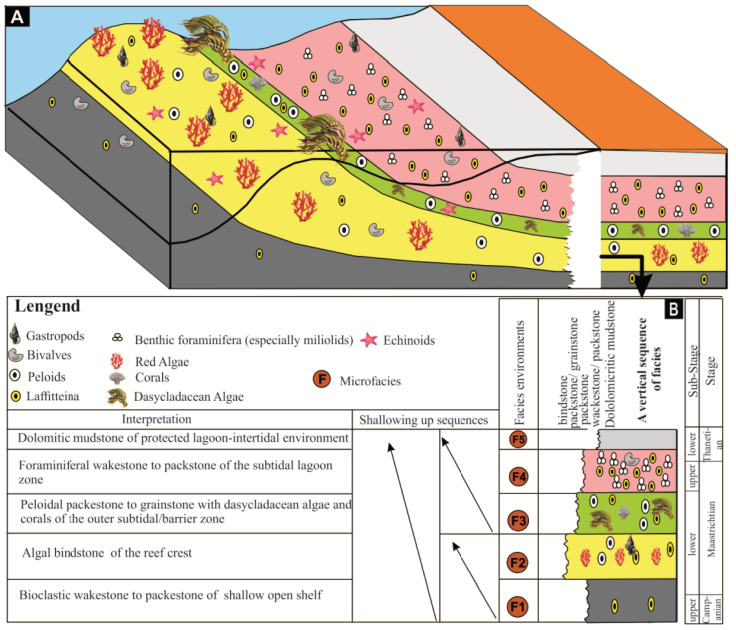
The first microfacies (F1) corresponds to bioclastic wackestone to packstone texture. This association consists of Laffitteina and miliolids with very little diversity, and a micritic matrix. The wackestone/ packestone texture and the micritc matrix show a generalized low energy of a circalittoral environment. This facies is comparable to SMF2 of Wilson (1975).
The low abundance of Laffitteina indicates a progressive facies change and the disappearance of the stress conditions that controlled the proliferation of Laffitteina.
The second microfacies (F2) of algal bindstone texture, which is a modification proposed by Embry and Klovan (1971) for the Dunham (1962) Boundstone type of limestone where the framework organisms were encrusting and binding the sediment. These red algae limestones show Sporolithon (Archaeolithothamnium sp.) horizons containing scattered bioclasts of bryozoans, gastropods, ostracods, and foraminifera that probably formed within the reef ridge (barrier) of a very shallow and high-energy water environment. This facies is comparable to SMF7 of Wilson (1975).
The microfacies (F3) corresponds to peloidal packstone to grainstone texture. This microfacies is mainly composed of non-skeletal grains (peloids) and some scattered bioclasts (notably Dasycladacean algae and corals). Typically, peloids and bioclastic shoals separate restricted lagoonal environments from deeper ramp environments (Flügel, 2010) and may act as broad and extensive barriers to currents and waves (Aghaei, 2012). This microfacies is comparable to SMF11 and SMF 12 described by Wilson (1975) indicating a shoal environment.
The fourth micofacies (F4) with foraminiferal wackestone to packstone texture correspond to Laffitteina and miliolids limestone. The low diversity of the other components (very rare echinoids and bryozoans) indicate that these sediments were deposited in protected, restricted, well-oxygenated and low-energy environments within the subtidal lagoon zone (Flügel, 2010; Wilson 1975), while the abundance of Laffitteina and miliolids indicates their tolerance to stressful conditions related especially to salinity variations and/or poor water circulation. This microfacies is very comparable to SMF18 described by Wilson (1975).
The fifth microfacies (F5) of dolomitic mud-stone texture corresponds to facies SMF 23 of Wilson (1975). The dolomitization affecting the limestones reflects an almost complete absence of fauna, except for some echinoids phantom. Such a sediment may be a reflection of a very shallow and protected lagoon-intertidal like environment.
In the Aurès Basin, the microfacies analysis of the Upper Cretaceous-Lower Paleogene suggests a shallow rimmed shelves carbonate platform with sequential organization mainly controlled by sea-level fluctuations (Figure 9A). Interpreting the observed changes in relative sea-level in this area reflects significant facies changes from a shallow open shelf environment at the base to shallow lagoon environments at the top of a shallowing-up sequence (Figure 9B). These data can be linked to a global cooling of the planet, affecting land and oceans at the K/Pg transition. This global cooling is not without affecting the eustatism, which designates the oscillations of the sea-level, susceptible to make regress by contraction of the volume of oceanic waters. These 2 regressions (temperature and eustatism) at the end of the Mesozoic, probably increased the conditions of “biological stress” (Li and Keller, 1998, Li et al., 2000) which caused a massacre of the benthic fauna. The effects of marine regression have been observed in North America, Paris Basin (Courtillot, 1995) and in the Aurès Basin, through the appearance of gaps at the base of the Paleogene.
7. Conclusion
The following conclusions can be drawn from this study:
1. Laffitteina mengaudi (Astre), L. aff. Mengaudi (Astre), L. conica Drooger, L. monodi Marie, L. bibensis Marie, L. oeztuerki Inan, L. marsicana Farinacci, L. erki (Sirel) and L. boluensis Dizer species are present in Algeria (Aurès Basin).
2. The species of Laffitteina genus in the Aurès Basin have been reported from the middle of the lower Maastrichtian to the lower Thanetian.
3. Laffitteina species can be used as index fossils to recognize the K/ Pg transition in the Aurès basin. The last occurrence of Laffitteina oztuerki marks the end of the Maastrichtian (Inan, 2004). It was found in its upper part of distribution, in association with Omphalocyclus macroporus (Lamarck), giving the age of the upper Maastrichtian. The first occurrence of Laffitteina erki marks the Thanetian age, it was found in association with Valvulina, which has never been mentioned in Cretaceous rocks.
Due to the absence of Danian and Selandian deposits (Belkhodja et al., 2004) after general emersion during the lower Paleocene (Marzoqi and Pascal, 2000), the Thanetian rocks are concordant with Upper Cretaceous strata indicating perhaps a parallel unconformity. It could therefore be proposed that the K/Pg transition occurs in the last 10 m of the massive limestone of the Ncham Member, very low where Belkhoudja and Bignot drew the boundary Beyond this, it is proposed that the beginning of the Paleocene transgression is started at least in the middle Thanetian.
Conditions, which may have been determined by salinity variations and/or poor water circulation.
4. Analysis of microfacies and benthic fora-miniferal assemblages suggests a shallow rimmed shelves carbonate with a shallowing-up sequential organization, primarily controlled by sea-level fluctuations.

